|
Research Ideas and Outcomes : Grant Proposal
|
|
Corresponding author: Malte Jochum (malte.jochum@idiv.de)
Received: 15 Mar 2019 | Published: 01 Apr 2019
© 2019 Nico Eisenhauer, Olga Ferlian, Dylan Craven, Jes Hines, Malte Jochum
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Eisenhauer N, Ferlian O, Craven D, Hines J, Jochum M (2019) Ecosystem responses to exotic earthworm invasion in northern North American forests. Research Ideas and Outcomes 5: e34564. https://doi.org/10.3897/rio.5.e34564
|
|
Abstract
Earth is experiencing a substantial loss of biodiversity at the global scale, while both species gains and losses are occurring at local and regional scales. The influence of these nonrandom changes in species distributions could profoundly affect the functioning of ecosystems and the essential services that they provide. However, few experimental tests have been conducted examining the influence of species invasions on ecosystem functioning. Even fewer have been conducted using invasive ecosystem engineers, which can have disproportionately strong influence on native ecosystems relative to their own biomass. The invasion of exotic earthworms is a prime example of an ecosystem engineer that is influencing many ecosystems around the world. In particular, European earthworm invasions of northern North American forests cause simultaneous species gains and losses with significant consequences for essential ecosystem processes like nutrient cycling and crucial services to humanity like soil erosion control and carbon sequestration. Exotic earthworms are expected to select for specific traits in communities of soil microorganisms (fast-growing bacteria species), soil fauna (promoting the bacterial energy channel), and plants (graminoids) through direct and indirect effects. This will accelerate some ecosystem processes and decelerate others, fundamentally altering how invaded forests function. This project aims to investigate ecosystem responses of northern North American forests to earthworm invasion. Using a novel, synthetic combination of field observations, field experiments, lab experiments, and meta-analyses, the proposed work will be the first systematic examination of earthworm effects on (1) plant communities and (2) soil food webs and processes. Further, (3) effects of a changing climate (warming and reduced summer precipitation) on earthworm performance will be investigated in a unique field experiment designed to predict the future spread and consequences of earthworm invasion in North America. By assessing the soil chemical and physical properties as well as the taxonomic (e.g., by the latest next-generation sequencing techniques) and functional composition of plant, soil microbial and animal communities and the processes they drive in four forests, work packages I-III take complementary approaches to derive a comprehensive and generalizable picture of how ecosystems change in response to earthworm invasion. Finally, in work package IV, meta-analyses will be used to integrate the information from work packages I-III and existing literature to investigate if earthworms cause invasion waves, invasion meltdowns, habitat homogenization, and ecosystem state shifts. Global data will be synthesized to test if the relative magnitude of effects differs from place to place depending on the functional dissimilarity between native soil fauna and exotic earthworms. Moving from local to global scale, the present proposal examines the influence of earthworm invasions on biodiversity–ecosystem functioning relationships from an aboveground–belowground perspective in natural settings. This approach is highly innovative as it utilizes the invasion by exotic earthworms as an exciting model system that links invasion biology with trait-based community ecology, global change research, and ecosystem ecology, pioneering a new generation of biodiversity–ecosystem functioning research.
Keywords
Aboveground-belowground interactions, biodiversity change, biodiversity-ecosystem functioning, earthworms, invasion, Lumbricidae, plant communities, soil food webs, global change
List of participants
Lee Frelich, Edward Johnson, Peter Reich, Tesfaye Wubet
State-of-the-art and objectives
Background
Anthropogenic activities cause species gains and losses at local and regional scales, although the functional consequences of those two simultaneous processes have been studied mostly in isolation from each other (
Native biodiversity and functions are particularly threatened in boreal and northern temperate forest ecosystems, where biological invasions were recently shown to represent the main drivers of biodiversity loss (
Earthworms as Invaders of northern North American Forests
Large parts of North America are lacking native earthworm species (
Earthworms are a major component of many terrestrial ecosystems (
Drivers of Earthworm Invasion
The presence of nonnative earthworms is strongly associated with human activities and infrastructure, such as timber harvesting, forest roads, cabins, and fishing areas (
Consequences of Earthworm Invasion
Northern temperate and boreal forests in North America developed over most of the past 10,000 years, since the last Pleistocene glaciation, in the absence of earthworms (
The presence of exotic earthworm species is likely to have profound effects on the native flora and fauna (Fig.

Extreme example of the potential consequences of earthworm invasion. Shown is a maple forest in Minnesota, USA, which has not yet been invaded by exotic earthworms (top), and another maple forest in that region that has been invaded by European earthworms (bottom). Please note that direct comparisons of those two forest stands are complicated and it is hard to identify cause and effect. Experimental manipulations are needed to study earthworm effects. Photo credit: Olga Ferlian (top) and Ulrich Pruschitzki (bottom).
Observational studies suggest that the invasion by exotic earthworms paves the way for subsequent invasions of plants and other organisms (
In preparation for this proposal, we established a database of previously published work to synthesize existing data on the effects of exotic earthworms on the aboveground and belowground biodiversity of northern North American deciduous forests (see Methodology for details). A recent meta-analysis based on this database found that earthworm ecological group richness, density, and biomass increase the cover of graminoid plants, while herbaceous and woody plants do not change in cover (
Although we found a significant reduction in soil invertebrate diversity in the presence of exotic endogeic earthworms, the data coverage remains insufficient to derive information on the trait composition and functional alterations of soil food webs. Results from a global change experiment in Germany (Hohenheim Climate Change Experiment [HoCC]), however, suggest major impacts of earthworms on the topology of soil food webs with effects depending on soil temperature (
The responses of ecosystem functions and services to earthworm invasion have rarely been assessed thus far. The most commonly reported process is litter decomposition, which is typically considerably increased by the presence of earthworms (
Consequences of Soil Biodiversity Loss and Compositional Shifts
Invasive earthworms cause a significant decline in plant and soil biodiversity (
Information on exotic earthworm effects on soil microbial communities is scarce, while several studies reported responses of total soil microbial biomass (
Conceptual Backbone and Hypotheses
Earthworms invade ecosystems in dynamic waves, inducing nonlinear alterations of communities and processes (
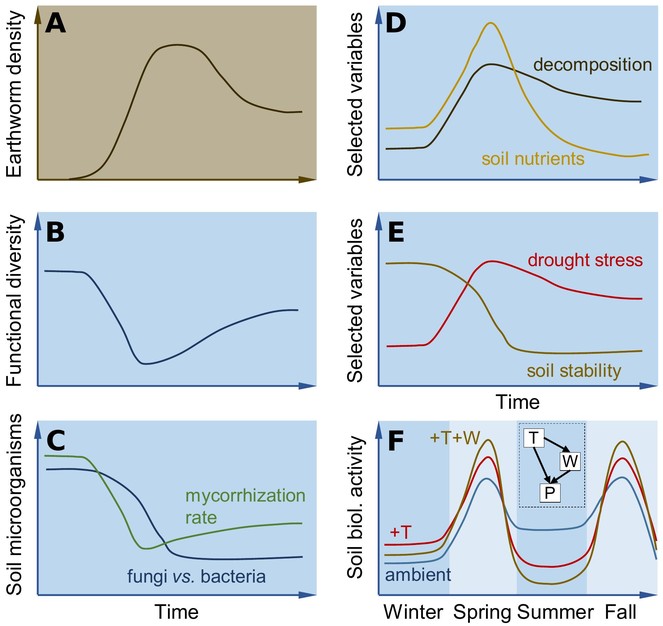
Conceptual depiction of the main hypotheses. [A] Earthworms invade ecosystems in waves (see text). Those dynamics induce nonlinear alterations of communities and processes. We expect to observe a decline in the functional diversity of plant and soil communities [B], recovering a bit after the invasion wave. We expect to see a shift in soil energy channels from a more fungal to a bacterial- dominated system with lower mycorrhization rates [C]. Further, decomposition [D] and drought stress are hypothesized to increase and soil stability to decrease [E]. Earthworms (W) and warming (T) will interact in intensifying seasonal dynamics in soil biological acitvity [F]. The inset indicates a path analysis differentiating direct effects of T on processes (P) from indirect effects through changes in W. See text for details.
Shifts in the plant community can be due to various mechanisms (H4) including changes in soil microbial community composition, mutualistic interactions (Fig.
By selecting for particular traits of plants and soil organisms and by facilitating plant invasion, exotic earthworms may cause invasional meltdowns (H5;
Methodology
Field Observation (FO)
For the FO, three transects will be established perpendicular to the invasion front by setting up five plots per transect for i) noninvaded areas, ii) areas invaded by epigeic earthworm species only, and (iii) areas invaded by all three ecological earthworm groups (Fig.
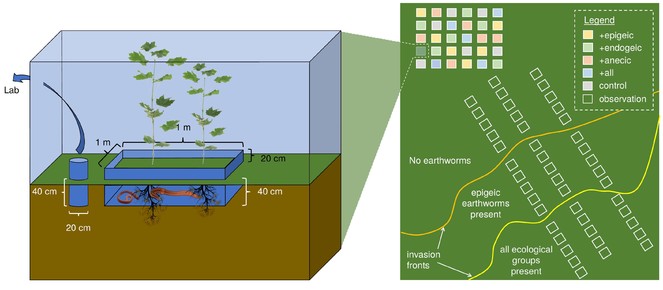
Experimental design. (Right panel) One forest patch (four spatially independent patches in total) with invasion fronts of epigeic earthworms (orange) and endogeic and anecic earthworms (yellow) present. Three transects à 15 observation plots (1 x 1 m;
The two aspen forests are located ~5 km apart in Kananaskis Valley in the front range of the Canadian Rocky Mountains in southwest Alberta at about 1410 m above sea level. The dominating tree species in both forests is trembling aspen (Populus tremuloides) interspersed with balsam poplar (Populus balsamifera). The understory is dense and mainly consists of herbs (see
All four study sites were used in previously published studies of European earthworm invasion impacts on native plant community structure and some groups of soil organisms, and have clearly documented leading edges of earthworm invasion (
Field Experiment (FE)
In the area without earthworms, a novel, systematic field experiment will be set up with experimental plots of 1 x 1 m in all four forests mentioned above (Fig.
Lab Experiment
Following the design of the FE, 30 intact soil cores (diameter 20 cm, depth 40 cm) will be excavated in noninvaded areas of each forest and brought to the laboratory (University of Calgary for sites in Alberta and University of Minnesota for the sites in Minnesota and Wisconsin). The same earthworm treatments and the same earthworm densities per m² will be established as in the field. Afterwards, 10 common native and 10 common exotic and invasive plant species will be added as seeds as done before (
Work Package I (WPI): Soil Food Webs and Processes
In each observational and experimental plot, yearly measurements of soil structure, soil pH, C and N concentrations (
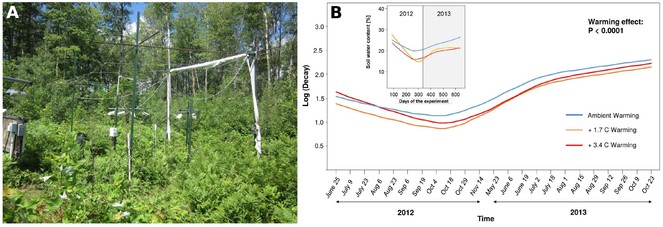
Exemplary plot of the B4WarmED experiment (Boreal Forest Warming in an Ecotone in Danger) in Cloquet, MN, USA [A] (
Data on the taxonomic composition and the topological structure of soil food webs (
Phenology of soil animal activity will be assessed via repeated measurements of bait lamina stripes. Preliminary tests in preparation for this proposal have shown that this approach is very powerful in capturing experimental treatment effects (soil warming) on the phenology of soil animal activity (Fig.
WPI will enable us to test hypotheses 1-3, 6-9, and to write at least five papers in international peer-reviewed journals on i) the responses of soil microbial communities and processes, ii) the topological structure of soil food webs, iii) the methodological approach of combining classical methods in the determination of density/biomass of soil organisms with cutting-edge sequencing techniques, iv) the phenology of soil biological activity, and v) soil BEF relationships. Multivariate statistical approaches, such as structural equation modeling, will facilitate the investigation of effects of different correlated explanatory variables to test multiple conflicting/parallel hypotheses (e.g.,
Work Package II (WPII): Plant Communities
In each observational and experimental plot, we will perform yearly (August) assessments of understory vegetation composition (plant species-specific cover;
The phytometers will allow standardized and comparable analyses: we will record signs of tree uprooting and growth responses. Yearly measurements of N and C:N in leaf tissue will serve as a proxy for N availability for plants, and δ13C will be measured to infer drought stress caused by earthworms (
The seed addition experiment in the laboratory allows investigating if exotic earthworms influence plant germination and establishment in intact plant communities (
WPII will test hypotheses 1, 4-9, resulting in at least five papers in peer-reviewed journals on i) the taxonomic and functional changes of plant communities along earthworm invasion fronts, ii) plant community responses to the presence of three different earthworm ecological groups, iii) the performance of tree saplings, iv) the germination and establishment of native and exotic plant species, and v) BEF relationships. For statistical analyses see WPI. For the analysis of BEF relationships it will be important to separate plant and soil diversity effects from other earthworm effects (e.g., changes in soil structure); therefore, earthworm density and biomass will be used as explanatory variables in structural equation models (
Work Package III (WPIII): Invasive Earthworms in a Changing World
The study will be conducted in the temperate-boreal forest ecotone of Minnesota, USA, in the framework of the B4WarmED experiment (Boreal Forest Warming at an Ecotone in Danger;
Earthworms are likely to intensify effects of warming and reduced precipitation on plant and soil communities and processes (Fig.
Structural equation modeling will allow investigating effects of warming and reduced precipitation on soil communities and activities by considering the responses and effects of exotic earthworm communities (Fig.
Work Package IV: Synthesis
In preparation of this proposal, we started creating a database of the effects of exotic earthworm invasion in North American forests on understory plant, soil invertebrate, and soil microbial communities by performing a search in the ISI Web of Science database using the keywords (‘earthworm*’) AND (‘exotic’ OR ‘invasive’). These words were selected in order to include a wide array of studies addressing the effects of exotic earthworms on above- and belowground communities. From the initial list of 359 studies, we examined each title and abstract to determine if they met the inclusion criteria. Our inclusion criteria were: i) that the study was performed in a North American forest ecosystem, ii) reported density, biomass, or presence/absence of exotic earthworms (identified to species or earthworm ecological group), iii) reported the abundance of plant species or plant functional groups in the forest understory, iv) reported the diversity and composition of soil invertebrate and soil microbial communities. Subsequently, we communicated directly with authors of the selected articles to obtain raw data. Thus far, we identified 24 unique studies meeting our inclusion criteria with 567 observations (12 studies) for plant communities, 724 observations (13 studies) for abundance of plant growth forms, 15 observations (7 studies) for richness of soil invertebrate communities, and 8 observations (6 studies) for abundance of soil microbial communities (Fig.
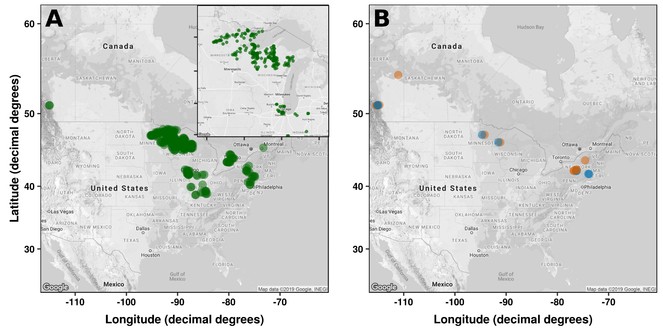
Location of study sites used in the meta-analyses to evaluate impacts of exotic earthworms on [A] plant and [B] soil invertebrate and microbial communities in North American forests. Each point corresponds to study sites (particular studies had multiple sites). Green: plant communities, orange: soil invertebrates, blue: soil microbial biomass.
This database will be extended by contacting researchers and performing regular literature searches. The iDiv Biodiversity Informatics Platform will host the database and guarantee standardized data publication. Moreover, a workshop will be organized at the German Centre for Integrative Biodiversity Research in Leipzig, Germany (http://www.idiv.de/en/idiv-global) to assemble comprehensive synthesis datasets on the consequences of exotic earthworm invasion with the help of international scientists. While the observations and experiments in this proposal focus on North America, the workshop will address earthworm invasion as a global phenomenon (
Synthesis work will investigate the overarching questions if exotic earthworm invasion i) causes habitat and community homogenization (
Acknowledgements
N.E. thanks seven anonymous reviewers for their evaluations of this proposal and acknowledges funding by the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement no. 677232). Moreover, we acknowledge funding by the German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig.
We acknowledge support from the German Research Foundation (DFG) and Leipzig University within the program of Open Access Publishing.
Funding program
The European Union’s Horizon 2020 research and innovation programme (grant agreement no. 677232).
Grant title
Ecosystem responses to exotic earthworm invasion in northern North American forests
Hosting institution
Institute of Biology, Leipzig University, Germany; German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Germany
References
-
Distribution and impacts of invasive earthworms in Canadian forest ecosystems.Biological Invasions11(1):59‑79. https://doi.org/10.1007/s10530-008-9320-4
-
Belowground biodiversity and ecosystem functioning.Nature515(7528):505‑511. https://doi.org/10.1038/nature13855
-
Historical anthropogenic disturbances influence patterns of non-native earthworm and plant invasions in a temperate primary forest.Biological Invasions17(4):1267‑1281. https://doi.org/10.1007/s10530-014-0794-y
-
Non-native invasive earthworms as agents of change in northern temperate forests.Frontiers in Ecology and the Environment2(8):427‑435. https://doi.org/10.2307/3868431
-
Strategies lombriciennes.Ecological Bulletins25:122‑132.
-
How do earthworms affect microfloral and faunal community diversity?Plant and Soil170(1):209‑231. https://doi.org/10.1007/bf02183068
-
Invasion by exotic earthworms alters biodiversity and communities of litter- and soil-dwelling oribatid mites.Diversity3(1):155‑175. https://doi.org/10.3390/d3010155
-
Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms.The ISME Journal6(8):1621‑1624. https://doi.org/10.1038/ismej.2012.8
-
Biodiversity loss and its impact on humanity.Nature486:59‑67. https://doi.org/10.1038/nature11148
-
Nematode functional guilds, not trophic groups, reflect shifts in soil food webs and processes in response to interacting global change factors.Pedobiologia58:23‑32. https://doi.org/10.1016/j.pedobi.2015.01.001
-
The unseen invaders: introduced earthworms as drivers of change in plant communities in North American forests (a meta-analysis).Global Change Biology23(3):1065‑1074. https://doi.org/10.1111/gcb.13446
-
Factors affecting the abundance of earthworms in soils. In: Edwards CA (Ed.)Earthworm Ecology.2nd edition.CRC Press,Boca Raton,91-114pp.
-
Of size and abundance.Nature351:268‑269. https://doi.org/10.1038/351268a0
-
Earthworm invasion, white-tailed deer and seedling establishment in deciduous forests of northeastern North America.Journal of Ecology103(1):153‑64. https://doi.org/10.1111/1365-2745.12350
-
Does global change increase the success of biological invaders?Trends in Ecology & Evolution14(4):135‑139. https://doi.org/10.1016/s0169-5347(98)01554-7
-
Plant diversity impacts decomposition and herbivory via changes in aboveground arthropods.PLoS ONE9(9):e106529. https://doi.org/10.1371/journal.pone.0106529
-
Earthworm ecology.CRC Presshttps://doi.org/10.1201/9781420039719
-
Invasion of a deciduous forest by earthworms: Changes in soil chemistry, microflora, microarthropods and vegetation.Soil Biology and Biochemistry39(5):1099‑1110. https://doi.org/10.1016/j.soilbio.2006.12.019
-
Earthworms as drivers of the competition between grasses and legumes.Soil Biology and Biochemistry40(10):2650‑2659. https://doi.org/10.1016/j.soilbio.2008.07.010
-
Animal ecosystem engineers modulate the diversity-invasibility relationship.PLoS ONE3(10):e3489. https://doi.org/10.1371/journal.pone.0003489
-
Exotic ecosystem engineers change the emergence of plants from the seed bank of a deciduous forest.Ecosystems12(6):1008‑1016. https://doi.org/10.1007/s10021-009-9275-z
-
Earthworm and belowground competition effects on plant productivity in a plant diversity gradient.Oecologia161(2):291‑301. https://doi.org/10.1007/s00442-009-1374-1
-
The action of an animal ecosystem engineer: Identification of the main mechanisms of earthworm impacts on soil microarthropods.Pedobiologia53(6):343‑352. https://doi.org/10.1016/j.pedobi.2010.04.003
-
Earthworms as seedling predators: Importance of seeds and seedlings for earthworm nutrition.Soil Biology and Biochemistry42(8):1245‑1252. https://doi.org/10.1016/j.soilbio.2010.04.012
-
Plant diversity effects on soil microorganisms support the singular hypothesis.Ecology91(2):485‑496. https://doi.org/10.1890/08-2338.1
-
The wave towards a new steady state: effects of earthworm invasion on soil microbial functions.Biological Invasions13(10):2191‑2196. https://doi.org/10.1007/s10530-011-0053-4
-
Decomposer diversity and identity influence plant diversity effects on ecosystem functioning.Ecology93(10):2227‑2240. https://doi.org/10.1890/11-2266.1
-
Interactive effects of global warming and ‘global worming’ on the initial establishment of native and exotic herbaceous plant species.Oikos121(7):1121‑1133. https://doi.org/10.1111/j.1600-0706.2011.19807.x
-
Global change belowground: impacts of elevated CO2, nitrogen, and summer drought on soil food webs and biodiversity.Global Change Biology18(2):435‑447. https://doi.org/10.1111/j.1365-2486.2011.02555.x
-
Plant diversity effects on soil food webs are stronger than those of elevated CO2 and N deposition in a long-term grassland experiment.Proceedings of the National Academy of Sciences110(17):6889‑6894. https://doi.org/10.1073/pnas.1217382110
-
Warming shifts ‘worming’: effects of experimental warming on invasive earthworms in northern North America.Scientific Reports4(1). https://doi.org/10.1038/srep06890
-
Organic textile dye improves the visual assessment of the bait-lamina test.Applied Soil Ecology82:78‑81. https://doi.org/10.1016/j.apsoil.2014.05.008
-
Invasive earthworms erode soil biodiversity: A meta-analysis.Journal of Animal Ecology87(1):162‑172. https://doi.org/10.1111/1365-2656.12746
-
Linking direct and indirect pathways mediating earthworms, deer, and understory composition in Great Lakes forests.Biological Invasions15(5):1057‑1066. https://doi.org/10.1007/s10530-012-0350-6
-
Importance of earthworm–seed interactions for the composition and structure of plant communities: A review.Acta Oecologica37(6):594‑603. https://doi.org/10.1016/j.actao.2011.03.001
-
Will environmental changes reinforce the impact of global warming on the prairie–forest border of central North America?Frontiers in Ecology and the Environment8(7):371‑378. https://doi.org/10.1890/080191
-
Trophic cascades, invasive species and body-size hierarchies interactively modulate climate change responses of ecotonal temperate-boreal forest.Philosophical Transactions of the Royal Society B: Biological Sciences367(1605):2955‑2961. https://doi.org/10.1098/rstb.2012.0235
-
Using earthworms as model organisms in the laboratory: Recommendations for experimental implementations.Pedobiologia53(2):119‑125. https://doi.org/10.1016/j.pedobi.2009.07.002
-
Restoration through reassembly: plant traits and invasion resistance.Trends in Ecology & Evolution23(12):695‑703. https://doi.org/10.1016/j.tree.2008.07.013
-
Exotic earthworm invasion and microbial biomass in temperate forest soils.Ecosystems7(1):45‑54. https://doi.org/10.1007/s10021-003-0129-9
-
Exotic European earthworm invasion dynamics in northern hardwood forests of Minnesota, USA.Ecological Applications15(3):848‑860. https://doi.org/10.1890/03-5345
-
Changes in hardwood forest understory plant communities in response to European earthworm invasions.Ecology87(7):1637‑1649. https://doi.org/10.1890/0012-9658(2006)87[1637:cihfup]2.0.co;2
-
Exotic earthworm effects on hardwood forest floor, nutrient availability and native plants: a mesocosm study.Oecologia155(3):509‑518. https://doi.org/10.1007/s00442-007-0925-6
-
Biodiversity effects on soil processes explained by interspecific functional dissimilarity.Science306(5698):1019‑1020. https://doi.org/10.1126/science.1101865
-
European buckthorn and Asian soybean aphid as components of an extensive invasional meltdown in North America.Biological Invasions12(9):2913‑2931. https://doi.org/10.1007/s10530-010-9736-5
-
Exotic earthworm invasions in North America: Ecological and policy implications.BioScience52(9):801‑811. https://doi.org/10.1641/0006-3568(2002)052[0801:eeiina]2.0.co;2
-
Pandora's box contained bait: The global problem of introduced earthworms.Annual Review of Ecology, Evolution, and Systematics39(1):593‑613. https://doi.org/10.1146/annurev.ecolsys.39.110707.173426
-
Effects of earthworm invasion on plant species richness in Northern Hardwood forests.Conservation Biology21(4):997‑1008. https://doi.org/10.1111/j.1523-1739.2007.00740.x
-
Invasion of exotic earthworms into North America and other regions.Earthworm Ecology441:75‑88. https://doi.org/10.1201/9781420039719.ch5
-
Organisms as ecosystem engineers.Ecosystem Management.Springer, New York
-
Soil invertebrates as ecosystem engineers: Intended and accidental effects on soil and feedback loops.Applied Soil Ecology32(2):153‑164. https://doi.org/10.1016/j.apsoil.2005.07.004
-
Genotypic richness and dissimilarity opposingly affect ecosystem functioning.Ecology Letters14(6):537‑545. https://doi.org/10.1111/j.1461-0248.2011.01613.x
-
Global climate change impacts in the United States.Cambridge University Press.
-
Influence of nonnative earthworms on mycorrhizal colonization of sugar maple (Acer saccharum).New Phytologist157(1):145‑153. https://doi.org/10.1046/j.1469-8137.2003.00649.x
-
Reduced density and nest survival of ground-nesting songbirds relative to earthworm invasions in Northern Hardwood forests.Conservation Biology25(5):983‑992. https://doi.org/10.1111/j.1523-1739.2011.01719.x
-
A simple and effective method to keep earthworms confined to open-top mesocosms.Applied Soil Ecology64:190‑193. https://doi.org/10.1016/j.apsoil.2012.12.008
-
Greenhouse-gas emissions from soils increased by earthworms.Nature Climate Change3:187‑194. https://doi.org/10.1038/nclimate1692
-
Earthworm invasions of ecosystems devoid of earthworms: effects on soil microbes.Biological Invasions8(6):1257‑1273. https://doi.org/10.1007/s10530-006-9020-x
-
The influence of invasive earthworms on indigenous fauna in ecosystems previously uninhabited by earthworms.Biological Invasions8(6):1275‑1285. https://doi.org/10.1007/s10530-006-9021-9
-
The effect of earthworm invasion on nutrient turnover, microorganisms and microarthropods in Canadian aspen forest soil.PhD Thesis, Darmstadt University of Technology
-
Functionally and phylogenetically diverse plant communities key to soil biota.Ecology94(8):1878‑1885. https://doi.org/10.1890/12-1936.1
-
A meta-analysis of declines in local species richness from human disturbances.Ecology and Evolution4(1):91‑103. https://doi.org/10.1002/ece3.909
-
Earthworm invasion as the driving force behind plant invasion and community change in Northeastern North American forests.Conservation Biology23(4):966‑974. https://doi.org/10.1111/j.1523-1739.2009.01168.x
-
Mechanisms linking plant community properties to soil aggregate stability in an experimental grassland plant diversity gradient.Plant and Soil373:285‑299. https://doi.org/10.1007/s11104-013-1791-0
-
A thesaurus for soil invertebrate trait-based approaches.PLoS ONE9(10):e108985. https://doi.org/10.1371/journal.pone.0108985
-
A novel photographic approach for monitoring the structural heterogeneity and diversity of grassland ecosystems.Journal of Plant Ecology7(6):518‑525. https://doi.org/10.1093/jpe/rtt065
-
Earthworms, as ecosystem engineers, influence multiple aspects of a salamander’s ecology.Oecologia165(3):745‑754. https://doi.org/10.1007/s00442-010-1775-1
-
Invasive Earthworms Deplete KeySoil Inorganic Nutrients (Ca, Mg, K,and P) in a Northern Hardwood forest.Ecosystems18:89‑102. https://doi.org/10.1007/s10021-014-9814-0
-
Invasive earthworms interact with abiotic conditions to influence the invasion of common buckthorn (Rhamnus cathartica).Oecologia178(1):219‑230. https://doi.org/10.1007/s00442-014-3175-4
-
Global biodiversity scenarios for the year 2100.Science (New York, N.Y.)287(5459):1770‑4. https://doi.org/10.1126/science.287.5459.1770
-
Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment.Nature468:553‑556. https://doi.org/10.1038/nature09492
-
Effects of earthworms on plant growth: patterns and perspectives.Pedobiologia47:846‑856. https://doi.org/10.1078/0031-4056-00270
-
Earthworm invasion alters enchytraeid community composition and individual biomass in northern hardwood forests of North America.Applied Soil Ecology83:159‑169. https://doi.org/10.1016/j.apsoil.2013.09.005
-
Earthworms modulate the effects of climate warming on the taxon richness of soilmeso- and macrofauna in an agricultural system.Agriculture Ecosystems and Environmenthttps://doi.org/10.1016/j.agee.2019.03.004
-
Plant diversity effects on soil microbial functions and enzymes are stronger than warming in a grassland experiment.Ecology96(1):99‑112. https://doi.org/10.1890/14-0088.1
-
Nonlinearity of effects of invasive ecosystem engineers on abiotic soil properties and soil biota.Oikos118(6):885‑896. https://doi.org/10.1111/j.1600-0706.2009.17405.x
-
Ecosystem effects of non-native earthworms in Mid-Atlantic deciduous forests.Biological Invasions13(5):1165‑1182. https://doi.org/10.1007/s10530-011-9959-0
-
Reduced feeding activity of soil detritivores under warmer and drier conditions.Nature Climate Change8(1):75‑78. https://doi.org/10.1038/s41558-017-0032-6
-
Nematode community shifts in response to experimental warming and canopy conditions are associated with plant community changes in the temperate-boreal forest ecotone.Oecologia175(2):713‑723. https://doi.org/10.1007/s00442-014-2927-5
-
Food webs: reconciling the structure and function of biodiversity.Trends in Ecology & Evolution27(12):689‑697. https://doi.org/10.1016/j.tree.2012.08.005
-
Biodiversity and ecosystem functioning.Annual Review of Ecology Evolution and Systematics45:471‑493. https://doi.org/10.1146/annurev-ecolsys-120213-091917
-
Invasion patterns of Lumbricidae into the previously earthworm-free areas of Northeastern Europe and the Western Great Lakes region of North America.Biological Invasions8(6):1223‑1234. https://doi.org/10.1007/s10530-006-9018-4
-
Ecological linkages between aboveground and belowground biota.Science304(5677):1629‑1633. https://doi.org/10.1126/science.1094875
-
Terrestrial ecosystem responses to species gains and ;osses.Science332(6035):1273‑1277. https://doi.org/10.1126/science.1197479
-
Terrestrial ecosystem responses to species gains and losses.Science (New York, N.Y.)332(6035):1273‑77. https://doi.org/10.1126/science.1197479
-
Resident plant diversity and introduced earthworms have contrasting effects on the success of invasive plants.Biological Invasions16(10):2181‑2193. https://doi.org/10.1007/s10530-014-0657-6
-
Differences in soil fungal communities between European beech (Fagus sylvatica L.) dominated forests are related to soil and understory vegetation.PLoS ONE7(10):e47500. https://doi.org/10.1371/journal.pone.0047500
-
Earthworms facilitate carbon sequestration through unequal amplification of carbon stabilization compared with mineralization.Nature Communications4(1):2576. https://doi.org/10.1038/ncomms3576