|
Research Ideas and Outcomes : Research Article
|
|
Corresponding author: Kiran Liversage (kiran.liversage@ut.ee)
Received: 14 Sep 2018 | Published: 11 Oct 2018
© 2018 Kiran Liversage, Maria Byrne
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Liversage K, Byrne M (2018) A note on life-history traits and conservation concerns for viviparous Australian seastars (Parvulastra parvivipara and P. vivipara). Research Ideas and Outcomes 4: e29766. https://doi.org/10.3897/rio.4.e29766
|
|
Abstract
The asterinid seastars Parvulastra parvivipara and P. vivipara share atypical viviparous reproductive modes that have made them of interest for research on life-history evolution and population genetics. This article briefly reviews life-history traits of these endemic Australian species and information on distribution and rarity, as well as providing some additional new analysis. Almost exclusive self-fertilisation has led to extreme genetic poverty in both species and viviparity limits dispersal potential causing relatively small geographical ranges. There is some evidence that the number of intertidal boulder-fields harbouring P. parvivipara, and the overall geographical range, may have become reduced in recent years. In addition, approximately 25 % of boulder-fields with P. parvivipara have been colonised by invasive oysters (Magallana gigas). To understand potential effects of oysters on P. parvivipara, we tested for correlations between P. parvivipara abundances and cover of oyster encrustations that included this invader (native + non-native oyster shells were assessed together because they produced similar encrustations and largely could not be differentiated). Linear regression showed no evidence, however, for any correlation. For P. vivipara, population survey data from the mid 1970’s to present shows marked decline in areas of previous abundance. Parallel with this decline, the boulder habitat has experienced infill and siltation and cementing of the substratum by M. gigas, sponges and other encrusters as well as anoxia. Important future research outcomes on consequences of atypical life-history traits may be allowed by continued research on these seastars but only if their populations are able to persist within the small number of boulder-fields where they occur. Current trends may indicate a need for conservation intervention.
Keywords
Intertidal boulder-field, boulder reef, rock pool, Crassostrea gigas, marine conservation
1. Life-history traits
Parvulastra parvivipara (Keough & Dartnall, 1978) and P. vivipara (Dartnall, 1969) are Australian small range endemic seastars occurring on coasts of western Eyre Peninsula (South Australia) and southeast Tasmania, respectively. Both species have atypical life-histories involving a diminutive size, simultaneous hermaphroditism, almost exclusive self-fertilisation, and brooding offspring in the gonads to an advanced juvenile which is achieved by sibling cannibalism (



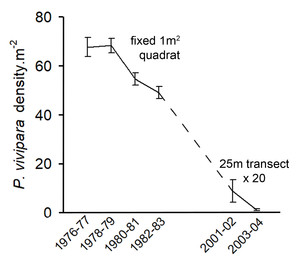
Photographs from P. parvivipara and P. vivipara habitat, and graph of P. vivipara population trends.
b: Photograph of a boulder underside in P. parvivipara habitat with extensive encrustation of oyster shells that includes invasive Pacific oysters (bar = 5 cm).
c: Photograph of P. vivipara during 1992 at Pit Water. Populations have become reduced in subsequent years which may be associated with increased siltation and overgrowth from encrusting species (bar = 5 cm).
d: Trends from the largest P. vivipara population at Pitt Water. Each point is the mean of adult densities from numerous sampling events over each 2 year period. During 1976-83 a fixed 1 m2 quadrat was sampled (
Both species use habitat underneath intertidal boulders, although P. vivipara is also associated with mussel beds. During night they emerge from under boulders (
2. Rarity
The overall distribution of P. parvivipara is limited to seven granitic headlands with the largest population occurring over 3924 m2 at the site “Smooth Pool” (
P. vivipara is known from six sites in southeast Tasmania (
3. Interactions with invasive species
Approximately 25 % of sites where P. parvivipara occur have been colonised by the non-native Pacific oyster Magallana gigas (Thunberg, 1793). M. gigas co-occurs with the seastar and other native oysters underneath boulders. One aim of this study was to use photographs of boulder undersurfaces in P. parvivipara habitat, taken by
The result showed that oyster shells reached covers up to 36 % (Fig.
Acknowledgements
We thank Lana Roediger for discussion on P. parvivipara biology.
Ethics and security
This project complies with the Estonian Code of Conduct for Research Integrity (http://www.eetika.ee/en/ethics-estonia/estonian-code-conduct-research-integrity).
Conflicts of interest
The authors declare no conflicts of interest.
References
-
Viviparity and intragonadal cannibalism in the diminutive sea stars Patiriella vivipara and P. parvivipara (family Asterinidae).Marine Biology125:551‑567. https://doi.org/10.1007/BF00353268
-
Causeway – Parvulastra vivipara population survival assessment.Ecomarine Pty Ltd, for the Department of Infrastructure, Energy and Resources, Tasmania,Hobart.
-
Shallow gene pools in the high intertidal: extreme loss of genetic diversity in viviparous sea stars (Parvulastra).Biology Letters9(5): . https://doi.org/10.1098/rsbl.2013.0551
-
A new species of viviparous Asterinid Asteroid from Eyre Peninsula, South Australia.Records of the South Australian Museum17:407‑416.
-
Habitat associations of a rare South Australian sea star (Parvulastra parvivipara) and a co-occurring chiton (Ischnochiton variegatus): implications for conservation.Pacific Conservation Biology21:234‑242. https://doi.org/10.1071/PC15017
-
What and when to eat? Investigating the feeding habits of an intertidal herbivorous starfish.Marine Biology163:1‑13. https://doi.org/10.1007/s00227-016-2942-7
-
The distribution and biology of Patiriella vivipara (Echinodermata: Asteroidea: Asterinidae) a sea star endemic to southeast Tasmania.Records of the Australian Museum50:161‑170. https://doi.org/10.3853/j.0067-1975.50.1998.1277
-
Updated information and previously unpublished observations on Patiriella vivipara, a seastar endemic to southeast Tasmania.The Tasmanian Naturalist123:24‑35.
-
Abundance and distribution of South Australia’s endemic sea star, Parvulastra parvivipara (Asteroidea: Asterinidae).Marine and Freshwater Research59:205‑213. https://doi.org/10.1071/MF07084