|
Research Ideas and Outcomes :
Grant Proposal
|
|
Corresponding author: Olga Ferlian (olga.ferlian@idiv.de)
Received: 26 Apr 2022 | Published: 07 Jun 2022
© 2022 Nico Eisenhauer, Paola Bonfante, François Buscot, Simone Cesarz, Carlos Guerra, Anna Heintz-Buschart, Jes Hines, Guillaume Patoine, Matthias Rillig, Bernhard Schmid, Kris Verheyen, Christian Wirth, Olga Ferlian
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Eisenhauer N, Bonfante P, Buscot F, Cesarz S, Guerra CA, Heintz-Buschart A, Hines J, Patoine G, Rillig MC, Schmid B, Verheyen K, Wirth C, Ferlian O (2022) Biotic Interactions as Mediators of Context-Dependent Biodiversity-Ecosystem Functioning Relationships. Research Ideas and Outcomes 8: e85873. https://doi.org/10.3897/rio.8.e85873
|
|
Abstract
Biodiversity drives the maintenance and stability of ecosystem functioning as well as many of nature’s benefits to people, yet people cause substantial biodiversity change. Despite broad consensus about a positive relationship between biodiversity and ecosystem functioning (BEF), the underlying mechanisms and their context-dependencies are not well understood. This proposal, submitted to the European Research Council (ERC), aims at filling this knowledge gap by providing a novel conceptual framework for integrating biotic interactions across guilds of organisms, i.e. plants and mycorrhizal fungi, to explain the ecosystem consequences of biodiversity change. The overarching hypothesis is that EF increases when more tree species associate with functionally dissimilar mycorrhizal fungi. Taking a whole-ecosystem perspective, we propose to explore the role of tree-mycorrhiza interactions in driving BEF across environmental contexts and how this relates to nutrient dynamics. Given the significant role that mycorrhizae play in soil nutrient and water uptake, BEF relationships will be investigated under normal and drought conditions. Resulting ecosystem consequences will be explored by studying main energy channels and ecosystem multifunctionality using food web energy fluxes and by assessing carbon storage. Synthesising drivers of biotic interactions will allow us to understand context-dependent BEF relationships. This interdisciplinary and integrative project spans the whole gradient from local-scale process assessments to global relationships by building on unique experimental infrastructures like the MyDiv Experiment, iDiv Ecotron and the global network TreeDivNet, to link ecological mechanisms to reforestation initiatives. This innovative combination of basic scientific research with real-world interventions links trait-based community ecology, global change research and ecosystem ecology, pioneering a new generation of BEF research and represents a significant step towards implementing BEF theory for human needs.
Keywords
biodiversity-ecosystem functioning, biodiversity effects, carbon sequestration, drought, energy flux, iDiv Ecotron, mycorrhiza, MyDiv, mutualists, nutrients, TreeDivNet
State-of-the-art and objectives
Biodiversity change and its consequences for ecosystems
Earth is experiencing a multitude of environmental changes that affect the composition and functioning of ecosystems. Concern that unprecedented rates of biodiversity change will alter ecosystem functioning and the provisioning of ecosystem services has prompted roughly three decades of research evaluating the relationship between biodiversity and ecosystem functioning (BEF) (
Ecological theory predicts that positive biodiversity effects on ecosystem functioning should arise if intraspecific competition in communities is higher than interspecific competition (
Above-Belowground Interactions in Biodiversity–Ecosystem Functioning Relationships
Aboveground-belowground interactions between plants and soil organisms may be particularly promising to explain plant BEF relationships and their context dependency (
Mycorrhizae – a Widespread Symbiosis Between Plants and Soil Fungi
Mycorrhizal fungi are a heterogeneous group of diverse fungal taxa, associated with the roots of > 90% of all plant species on Earth (
The Global Role of Mycorrhizal Fungi for Ecosystem Functioning
The global extent, distribution and functional composition of forests is central to our understanding of the functioning of the terrestrial biosphere (
Across these different biomes, AMF and EMF vegetation were shown to store substantial amounts of C in aboveground biomass (241 and 100 gigatons (GT), respectively; non-mycorrhizal vegetation: 29 ± 5.5 GT C only;
Most studies on the effects of AMF and EMF trees have been done in laboratory or nursery conditions (
Context-Dependent Biodiversity Effects on Ecosystem Functioning
Unlike most controlled BEF experiments, real landscapes are heterogeneous and exhibit strong gradients in abiotic conditions, such as precipitation regime or nutrient availability. To apply results from BEF experiments to natural landscapes and to develop biodiversity-based management applications, we need to understand the environmental and biotic context dependency of BEF relationships (
The Role of Mycorrhizal Fungi in BEF Relationships with Focus on Proposed Work
This section summarises important knowledge gaps and main promising research directions to link to the proposed work packages (WPs) that are expected to advance BEF research.
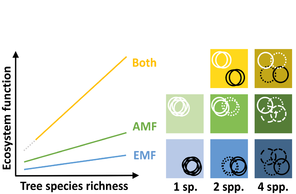
Similar to their plant partners, different mycorrhizal types and taxa have evolved ways to lower competition in space and time and possess various traits (
Resource acquisition by mycorrhizal fungi often targets plant-unavailable or limiting resources, such as N, P and water (
The diversity and type of mycorrhizal networks have been recognised as significant drivers of water uptake during extended droughts (
Biodiversity is a significant driver of ecosystem multiple ecosystem functions (
The Importance of Tree BEF Studies and Promising Links to Real-World Applications
To advance BEF theory, study mycorrhiza-mediated BEF mechanisms and to disentangle the effects of mycorrhizal types from that of plant species identity and other abiotic and biotic interactions, it is crucial to manipulate mycorrhizal types along a plant diversity gradient in experimental studies (
Main Objectives and Overarching Hypothesis
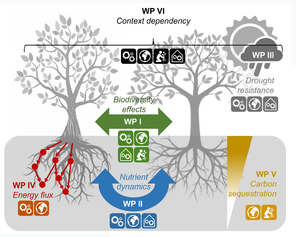
The functioning and service supply of ecosystems in the face of anthropogenic environmental and biodiversity change represents a cornerstone of ecological research and a pressing societal issue. Despite broad consensus about a positive BEF relationship, the underlying ecological mechanisms and their context-dependencies are not well understood. This proposal aims at filling this knowledge gap by providing a novel conceptual framework for integrating biotic interactions across guilds of organisms, more specifically between plants and mycorrhizal types, to explain resource-use complementarity in plants and its consequences for plant performance and community multifunctionality. Using a combination of field and mesocosm experiments, as well as meta-level analyses (Fig.
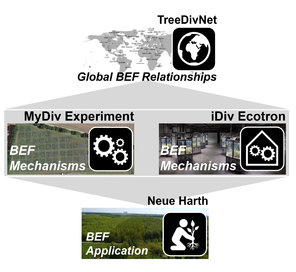
Structure, research platforms and work packages (WPs) of the proposed project. (A) WPs will work and collaborate in four main research platforms. Hypothesised mechanisms between biodiversity and ecosystem function (BEF) will be studied in the MyDiv Experiment and the iDiv Ecotron. While the MyDiv Experiment allows exploring BEF relationships under natural environmental conditions, the iDiv Ecotron enables studies under more controlled conditions and the test of additional drivers (e.g. nutrients, drought). The global experimental network TreeDivNet will allow us to generalise our findings and/or to study context-dependent BEF relationships. The local reforestation site Neue Harth will allow us to directly test the real-world, management-relevant implications of BEF relationships. Platforms are represented with different icons that are used in (B) to indicate which WP builds on these platforms. The six different WPs take complementary approaches to study important ecosystem components and facilitate synthesis.
Methodology
From Experiments to Real World: the Platforms Used across the Proposal
The MyDiv Experiment – a Field Experiment on the Role of Mycorrhizae in BEF
In 2015, the MyDiv Experiment (Fig.
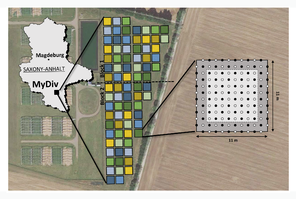
(A) Location of the MyDiv Experiment, experimental design with colour coding and within-plot experimental design with the core area (light grey) and planting pattern (
Notably, given the significant establishment effects of ecological experiments, including disturbances (
The iDiv Ecotron – an Experimental Facility to Study Multitrophic BEF
Recently, the iDiv Ecotron was set up to address the perpetual claim that BEF research in terrestrial ecosystems needs to move beyond the manipulation of diversity at single trophic levels to embrace the multitrophic complexity of ecological communities (



The iDiv Ecotron. (A) Photo showing the iDiv Ecotron facility with EcoUnits. (B) Technical drawing of an EcoUnit illustrating the lysimeter function with soil sensors and root scanners. (C) Photo of a preliminary experiment in preparation of this proposal with beech saplings. In the proposed experiments, we will use the same lysimeter set-up to be able to incubate up to 96 independent mesocosms. Figures were modified after
TreeDivNet – a Global Network of Tree Diversity Experiments
The global nature of environmental problems, such as climate change, desertification and biodiversity loss, requires the establishment of research approaches that, in most cases, exceed the capacity of single countries or research groups (
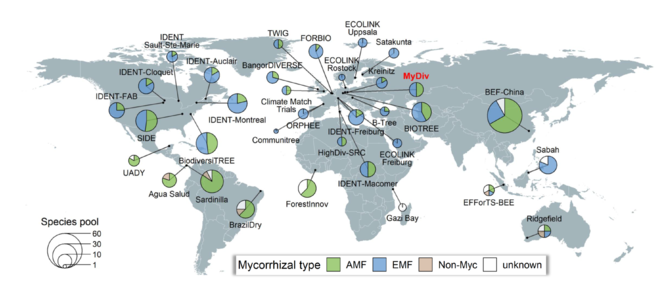
The global network of tree diversity experiments “TreeDivNet”. Each experimental site is represented by one pie chart (some experiments have multiple sites). The sizes of the pie charts correspond to the respective tree species pool of the experiment. Based on information from TRY (retrieved in December 2019;
Neue Harth – a Reforestation Initiative after Open Coal Mining
There are many human activities that threaten the biodiversity and functioning of ecosystems, but habitat destruction is one of the most pervasive ones (


Reforestation of destroyed ecosystems. (A) Consequences of open coal mining in the south of Leipzig, Germany (2009; photo: Ronny Schmidt, GeoWerkstatt Leipzig e.V.). Roughly 800 ha of mixed forest were destroyed between 1921 and 1999. (B) Reforested area close to Leipzig, Germany (2015; photo: N. Eisenhauer). Forest patches were planted as monocultures and mixed forests of different diversity levels and now allow the study of the ecosystem consequences of different management decisions.
We have been in contact with the forest managers of the Neue Harth, who strongly support scientific work in these reforested areas. Together, we were able to assess the extent, tree species identity and diversity of restored forest patches. In this area, we defined 18 independent ~ 20-year old forest stands (> 2 ha each), containing 10 different tree species associating with different mycorrhizal types (Acer campestre [AMF], Acer platanoides [AMF], Betula pendula [EMF], Fagus sylvatica [EMF], Robinia pseudoacacia [AMF; association with N-fixing rhizobacteria], Tilia cordata [EMF], Pinus sylvestris [EMF], Populus balsamifera [EMF], Quercus rubra [EMF] and Quercus robur [EMF]), ranging from monocultures to 6-species mixtures. This setting represents a real-world reforestation scenario with tree species of high local relevance. The two oak species (with Q. robur being the phytometer species in the MyDiv Experiment) occur in replicated monocultures (n = 2 for both species), 2-species mixtures (n = 3) and 5-species mixtures (n = 3), enabling studies on the ecosystem consequences of diversifying oak plantations both in terms of tree species richness and mycorrhizal type. Further, recently developed analytical methods will allow us to explore complementarity and selection effects with these data (
Work Packages
The proposed work is divided into six highly complementary work packages (WPs; addressing the six main objectives outlined above) that are further subdivided into 18 tasks (TAs) in total. Four WPs will be carried out by more experienced scientists (Prof. Dr. Nico Eisenhauer and three postdocs) and the other two WPs will be conducted by PhD students. Each scientific TA is supposed to deliver at least one publication in an international, peer-reviewed journal. Team members will have complementary expertise and work together in joint experiments, sampling campaigns and synthesis projects. WPs I and VI will provide the conceptual backbone of the project, integrate the information from all other WPs and facilitate collaboration through multiple workshops. This interdisciplinary and integrative project ranges from local-scale process studies to global syntheses, to link the main ecological mechanisms to reforestation initiatives. The interdisciplinary nature of the project will build bridges between microbial, plant and ecosystem ecology and between basic research and applied aspects related to the restoration of multifunctional forests.
WP I: Biodiversity effects
This WP will focus on the strength of BEF relationships and how net biodiversity effects (NBEs), complementarity effects (CEs) and selection effects (SEs) (based on the additive partitioning method;
This WP will support all other WPs by coordinating the planned work, providing baseline data for internal and external collaborators, as well as supervising and mentoring postdocs and PhD students (TA-1-1). To provide baseline data on root mycorrhization for all other WPs, we will analyse the diversity and colonisation rate of AMF and EMF in roots using microscopy and NGS (amplicon and shotgun sequencing) in collaboration with iDiv’s Metagenomics Support Unit (Prof. Dr. Francois Buscot and Dr. Anna Heintz-Buschart). We will analyse the mycorrhiza diversity in tree roots in the MyDiv Experiment and in four selected experiments in TreeDivNet. These additional experiments (IDENT-Montreal, IDENT-FAB, IDENT-Macomer, BEF China) were chosen based on the criteria that they: (i) have at least three AMF and three EMF tree species in their species pool, (ii) were set up > 5 years ago and (iii) have the same diversity levels as the MyDiv Experiment (1, 2 and 4 species).
Similar to what was done in 2017 (
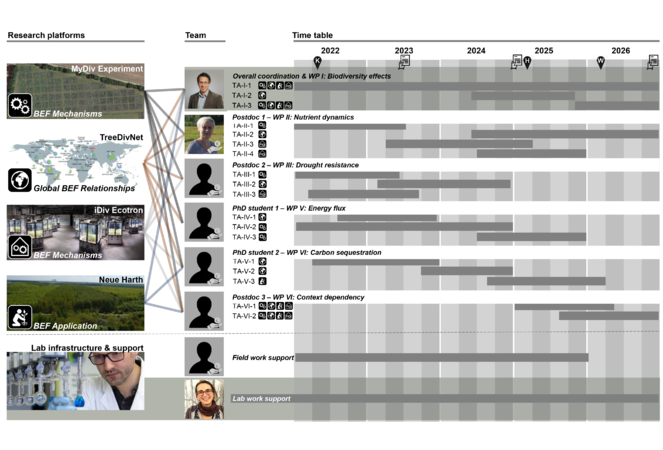
Timetable of the proposed project. Given are the main research platforms and the duration of each task (TA) within work packages (WPs). Pin icon: workshops; sheet icon: regular ERC reporting; K: Kick-off workshop, H: Hands-on workshop; W: Wrap-up workshop; hand icon: requested personnel. Icons on TAs indicate the research platform where the research will be conducted to illustrate the potential for collaboration.
Work package II: Nutrient dynamics
This WP will investigate the potential nutrient-related mechanisms underlying mycorrhiza-mediated tree diversity effects. Taxonomically and functionally diverse mycorrhizal communities may increase ecosystem functioning by enhancing the access and use of the available resource pool to plants resulting in relaxed plant-plant competition for soil resources (
In this WP, we will focus on soil nutrient dynamics as well as plant and soil stoichiometry as affected by tree diversity and mycorrhizal types. In TA-II-1, we will study C, N and P concentrations of leaves, soil and soil microbial biomass in the MyDiv Experiment. Briefly, as done in
Work package III: Drought resistance
This WP will investigate if mycorrhizal fungi can mitigate drought effects on trees. The increasing frequency and intensity of droughts is threatening the biodiversity and functioning of forest ecosystems (
In this WP, we will focus on tree growth, survival and physiology as important indicators of tree performance under drought conditions. In TA-III-1, we will analyse detailed tree inventory data from the MyDiv Experiment including the exceptionally dry summers in 2018 and 2019. We will analyse tree growth and mortality as well as δ13C in leaves as a proxy of stomatal conductance (as done in
Work package IV: Energy flux
This WP will investigate tree diversity and mycorrhiza effects on energy flux and ecosystem multifunctionality. Biodiversity is known to determine multitrophic energy use efficiency, flow and storage in grasslands (
In this WP, we will explore the structure and functioning of soil food webs. In TA-IV-1, we will collect soil from 12 selected long-term tree diversity experiments in TreeDivNet (Fig.
Work package V: Carbon sequestration
This WP will investigate tree diversity and mycorrhiza effects on soil C storage. Plant diversity has been shown to increase grassland soil C storage (
In this WP, we will investigate the drivers of soil C storage. In TA-V-1, we will collect soil from all 28 tree diversity experiments in TreeDivNet (Fig.
Work package VI: Context dependency
This WP will investigate the context dependency of BEF relationships (
In this WP, we will integrate work across all other WPs by investigating the context dependency of BEF relationships and the role of tree-mycorrhiza interactions explaining these differences. In TA-VI-1, we will build on an extensive literature review of empirical and theoretical work on this topic, as well as discussions in workshops (Fig.
Risks and Solutions
Large projects like the one proposed here are always associated with multiple potential risks that threaten the success of subprojects. However, we have ample experience with running and coordinating large experimental platforms like the iDiv Ecotron, as well as large research consortia with ~ 100 members like the Jena Experiment (
Ecological experiments can fail due to many logistic reasons. Here, we will avoid major risks by proposing cutting-edge research in well-established experiments. TreeDivNet is a well-functioning collaborative network; the MyDiv Experiment (since 2015;
The comparably young age of included experiments (< 20 y) could be criticised. While this is a common phenomenon in ecological experiments, including BEF (
Scientific and Broader Impact
Scientific impact. Each task is expected to deliver at least one scientific publication in leading multidisciplinary and disciplinary journals with major advances in ecosystem ecology and BEF. Moreover, we will make our data (e.g. mycorrhizal type, soil microbial and abiotic data) available to TreeDivNet (and beyond), which will facilitate the contribution to further syntheses. In addition, we will publish summaries of our main results in journals for foresters and land managers, such as that which has been done recently (
Outreach and societal impact. Since 2014, we have been very active in public outreach with 18 TV shows/interviews, 21 radio interviews, 189 print articles, 334 online articles and 55 other outreach activities, including public talks and demonstrations for politicians. In the framework of this proposal, it is planned to produce an educational video on ‘Human benefits of the tree-fungi friendship’ that provides information on the main outcomes of our research in collaboration with the YouTube Channel MinuteEarth. The applied aspects of this proposal related to reforestation are at the heart of current scientific debate and political agendas (e.g.
Open science. We are committed to open science and we will publish our findings in open-access journals, including underlying datasets and code, if applicable. This approach is supposed to fight the publication bias of under-represented, less well-funded areas of the globe and non-significant results. We strongly believe that our approach to fair open access publishing will serve as a role model, not only for soil ecology, but for scientific publishing in general.
Schedule and Intermediate Goals
The present project encompasses several independent sub-projects (Fig.
Acknowledgements
N.E. thanks eight anonymous reviewers for their evaluations of this proposal in the context of the Consolidator Grant application within the European Research Council (ERC). We thank Ulrich Brose and Malte Jochum for their valuable comments on earlier versions of this proposal. Moreover, we acknowledge funding by the German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig. We acknowledge support from the German Research Foundation (DFG).
Proposal title
Biotic Interactions as Mediators of Context-Dependent Biodiversity-Ecosystem Functioning Relationships – BIOCODE
Funding program
The European Research Council’s (ERC) Consolidator Grant 2021.
References
- A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests.Forest Ecology and Management259(4):660‑684. https://doi.org/10.1016/j.foreco.2009.09.001
- Consequences of widespread tree mortality triggered by drought and temperature stress.Nature Climate Change3(1):30‑36. https://doi.org/10.1038/nclimate1635
- Options of partners improve carbon for phosphorus trade in the arbuscular mycorrhizal mutualism.Ecology Letters19(6):648‑656. https://doi.org/10.1111/ele.12601
- Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis.Mycorrhiza11(1):3‑42. https://doi.org/10.1007/s005720100097
- Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage.Nature505(7484):543‑545. https://doi.org/10.1038/nature12901
- Global imprint of mycorrhizal fungi on whole-plant nutrient economics.Proceedings of the National Academy of Sciences116(46):23163‑23168. https://doi.org/10.1073/pnas.1906655116
- Biodiversity effects on ecosystem functioning respond unimodally to environmental stress.Ecology Letters21(8):1191‑1199. https://doi.org/10.1111/ele.13088
- Energy flux: the link between multitrophic biodiversity and ecosystem functioning.Trends in Ecology & Evolution33(3):186‑197. https://doi.org/10.1016/j.tree.2017.12.007
- Biodiversity enhances the multitrophic control of arthropod herbivory.Science Advances6(45):6603. https://doi.org/10.1126/sciadv.abb6603
- The Future of Complementarity: Disentangling Causes from Consequences.Trends in Ecology & Evolution34(2):167‑180. https://doi.org/10.1016/j.tree.2018.10.013
- Limited evidence for spatial resource partitioning across temperate grassland biodiversity experiments.Ecologye02905https://doi.org/10.1002/ecy.2905
- Positive interactions in communities.Trends in Ecology & Evolution9(5):191‑193. https://doi.org/10.1016/0169-5347(94)90088-4
- Rooting theories of plant community ecology in microbial interactions.Trends in Ecology & Evolution25(8):468‑478. https://doi.org/10.1016/j.tree.2010.05.004
- Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis.Nature Communications1(1):1‑11. https://doi.org/10.1038/ncomms1046
- Finding generality in ecology: a model for globally distributed experiments.Methods in Ecology and Evolution5(1):65‑73. https://doi.org/10.1111/2041-210X.12125
- Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil.Soil Biology and Biochemistry17(6):837‑842. https://doi.org/10.1016/0038-0717(85)90144-0
- Mycorrhizal associations and other means of nutrition of vascular plants: understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis.Plant and Soil320(1-2):37‑77. https://doi.org/10.1007/s11104-008-9877-9
- Evolutionary studies of ectomycorrhizal fungi: recent advances and future directions.Canadian Journal of Botany82(8):1122‑1132. https://doi.org/10.1139/b04-021
- Mycorrhizal type determines the magnitude and direction of root‐induced changes in decomposition in a temperate forest.New Phytologist206(4):1274‑1282. https://doi.org/10.1111/nph.13303
- Biodiversity increases multitrophic energy use efficiency, flow, and storage in grasslands.Nature Ecology & Evolution4(3):393‑405. https://doi.org/10.1038/s41559-020-1123-8
- The functional role of producer diversity in ecosystems.American Journal of Botany98(3):572‑592. https://doi.org/10.3732/ajb.1000364
- Biodiversity loss and its impact on humanity.Nature486(7401):59‑67. https://doi.org/10.1038/nature11148
- Tree diversity effects on soil microbial biomass and respiration are context dependent across forest diversity experiments.Global Ecology and Biogeography00:1‑14. https://doi.org/10.1111/geb.13461
- Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function.New Phytologist214(1):432‑442. https://doi.org/10.1111/nph.14343
- Differential soil fungus accumulation and density dependence of trees in a subtropical forest.Science366(6461):124‑128. https://doi.org/10.1126/science.aau1361
- Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees.Proceedings of the National Academy of Sciences113(31):8741‑8746. https://doi.org/10.1073/pnas.1601006113
- Biodiversity inhibits parasites: broad evidence for the dilution effect.Proceedings of the National Academy of Sciences112(28):8667‑8671. https://doi.org/10.1073/pnas.1506279112
- How to estimate complementarity and selection effects from an incomplete sample of species.Methods in Ecology and Evolution10(12):2141‑2152. https://doi.org/10.1111/2041-210X.13285
- The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts.Soil Biology and Biochemistry42(5):679‑698. https://doi.org/10.1016/j.soilbio.2009.12.006
- Tree mycorrhizal type predicts within‐site variability in the storage and distribution of soil organic matter.Global Change Biology24(8):3317‑3330. https://doi.org/10.1111/gcb.14132
- Multiple facets of biodiversity drive the diversity-stability relationship.Nature Ecology & Evolution2(10):1579‑1587. https://doi.org/10.1038/s41559-018-0647-7
- Evolution of interdisciplinarity in biodiversity science.Ecology and Evolution9(12):6744‑6755. https://doi.org/10.1002/ece3.5244
- Mapping tree density at a global scale.Nature525(7568):201‑205. https://doi.org/10.1038/nature14967
- Predicting ecosystem stability from community composition and biodiversity.Ecology Letters16(5):617‑625. https://doi.org/10.1111/ele.12088
- Pervasive human-driven decline of life on Earth points to the need for transformative change.Science366(6471). https://doi.org/10.1126/science.aax3100
- Biodiversity effects in the wild are common and as strong as key drivers of productivity.Nature549(7671):261‑264. https://doi.org/10.1038/nature23886
- Distinct seasonal assemblages of arbuscular mycorrhizal fungi revealed by massively parallel pyrosequencing.New Phytologist190(3):794‑804. https://doi.org/10.1111/j.1469-8137.2010.03636.x
- Earthworms as drivers of the competition between grasses and legumes.Soil Biology and Biochemistry40(10):2650‑2659. https://doi.org/10.1016/j.soilbio.2008.07.010
- Impacts of earthworms and arbuscular mycorrhizal fungi (Glomus intraradices) on plant performance are not interrelated.Soil Biology and Biochemistry41(3):561‑567. https://doi.org/10.1016/j.soilbio.2008.12.017
- Plant diversity effects on soil microorganisms support the singular hypothesis.Ecology91(2):485‑496. https://doi.org/10.1890/08-2338.1
- Aboveground-belowground interactions as a source of complementarity effects in biodiversity experiments.Plant and Soil351(1-2):1‑22. https://doi.org/10.1007/s11104-011-1027-0
- Decomposer diversity and identity influence plant diversity effects on ecosystem functioning.Ecology93(10):2227‑2240. https://doi.org/10.1890/11-2266.1
- Increasing plant diversity effects on productivity with time due to delayed soil biota effects on plants.Basic and Applied Ecology13(7):571‑578. https://doi.org/10.1016/j.baae.2012.09.002
- Plant diversity effects on soil food webs are stronger than those of elevated CO2 and N deposition in a long-term grassland experiment.Proceedings of the National Academy of Sciences110(17):6889‑6894. https://doi.org/10.1073/pnas.1217382110
- Biodiversity-ecosystem function experiments reveal the mechanisms underlying the consequences of biodiversity change in real world ecosystems.Journal of Vegetation Science27(5):1061‑1070. https://doi.org/10.1111/jvs.12435
- Root biomass and exudates link plant diversity with soil bacterial and fungal biomass.Scientific Reports7(1):1‑8. https://doi.org/10.1038/srep44641
- Aboveground-belowground interactions drive the relationship between plant diversity and ecosystem function.Research Ideas and Outcomes4:23688. https://doi.org/10.3897/rio.4.e23688
- Artenverlust schmälert Ökosystemleistungen.Ökologie & Landbau2|2018:50‑51.
- From climate chambers to biodiversity chambers.Frontiers in Ecology and the Environment16(3):136‑137. https://doi.org/10.1002/fee.1784
- Artenvielfalt verbessert die Funktionsweise von Ökosystemen.Bionachrichten, Ausgabe36‑37.
- The dark side of animal phenology.Trends in Ecology & Evolution33(12):898‑901. https://doi.org/10.1016/j.tree.2018.09.010
- Plant diversity maintains multiple soil functions in future environments.ELife7:41228. https://doi.org/10.7554/eLife.41228
- Biotic interactions, community assembly, and eco-evolutionary dynamics as drivers of long-term biodiversity-ecosystem functioning relationships.Research Ideas and Outcomes: The Open Science Journal5:47042. https://doi.org/10.3897/rio.5.e47042
- A multitrophic perspective on biodiversity-ecosystem functioning research.Advances in Ecological Research61:1. https://doi.org/10.1016/bs.aecr.2019.06.001
- Stepping up EU Action to Protect and Restore the World’s Forests.COM(2019)352:22.
- A new spectral similarity water index for the estimation of leaf water content from hyperspectral data of leaves.Remote Sensing of Environment196:13‑27. https://doi.org/10.1016/j.rse.2017.04.029
- Leaf and root C-to-N ratios are poor predictors of soil microbial biomass C and respiration across 32 tree species.Pedobiologia65:16‑23. https://doi.org/10.1016/j.pedobi.2017.06.005
- Growing Research Networks on Mycorrhizae for Mutual Benefits.Trends in Plant Science23(11):975‑984. https://doi.org/10.1016/j.tplants.2018.08.008
- Mycorrhiza in tree diversity–ecosystem function relationships: conceptual framework and experimental implementation.Ecosphere9(5). https://doi.org/10.1002/ecs2.2226
- Mycorrhizal fungi and their multifunctional roles.Mycologist18(2):91‑96. https://doi.org/10.1017/S0269915X04002058
- Plant species richness and functional groups have different effects on soil water content in a decade‐long grassland experiment.Journal of Ecology107(1):127‑141. https://doi.org/10.1111/1365-2745.13046
- Plant functional composition influences rates of soil carbon and nitrogen accumulation.Journal of Ecology96(2):314‑322. https://doi.org/10.1111/j.1365-2745.2007.01345.x
- fluxweb: An R package to easily estimate energy fluxes in food webs.Methods in Ecology and Evolution10(2):270‑279. https://doi.org/10.1111/2041-210X.13109
- Networked and embedded scientific experiments will improve restoration outcomes.Frontiers in Ecology and the Environment16(5):288‑294. https://doi.org/10.1002/fee.1810
- Quantifying resource use complementarity in grassland species: a comparison of different nutrient tracers.Pedobiologia57(4-6):251‑256. https://doi.org/10.1016/j.pedobi.2014.09.001
- Scaling‐up biodiversity‐ecosystem functioning research.Ecology Letters23(4):757‑776. https://doi.org/10.1111/ele.13456
- Integrative modelling reveals mechanisms linking productivity and plant species richness.Nature529(7586):390‑393. https://doi.org/10.1038/nature16524
- Tree diversity does not always improve resistance of forest ecosystems to drought.Proceedings of the National Academy of Sciences111(41):14812‑14815. https://doi.org/10.1073/pnas.1411970111
- Synthesis and future research directions linking tree diversity to growth, survival, and damage in a global network of tree diversity experiments.Environmental and Experimental Botany152:68‑89. https://doi.org/10.1016/j.envexpbot.2017.12.015
- Diversity-dependent temporal divergence of ecosystem functioning in experimental ecosystems.Nature Ecology & Evolution1(11):1639‑1642. https://doi.org/10.1038/s41559-017-0325-1
- Diversity‐dependent plant-soil feedbacks underlie long‐term plant diversity effects on primary productivity.Ecosphere10(4):02704. https://doi.org/10.1002/ecs2.2704
- Interspecific competition alters leaf stoichiometry in 20 grassland species.Oikos127(7):903‑914. https://doi.org/10.1111/oik.04907
- Mixing tree species associated with arbuscular or ectotrophic mycorrhizae reveals dual mycorrhization and interactive effects on the fungal partners.Ecology and Evolution00,1:17. https://doi.org/10.1002/ece3.7437
- Endogenous rhythmic growth, a trait suitable for the study of interplays between multitrophic interactions and tree development.Perspectives in Plant Ecology, Evolution and Systematics19:40‑48. https://doi.org/10.1016/j.ppees.2016.02.003
- Towards an integration of biodiversity-ecosystem functioning and food web theory to evaluate relationships between multiple ecosystem services.Advances in ecological research.53.Academic Presshttps://doi.org/10.1016/bs.aecr.2015.09.001
- A meta‐analysis of context‐dependency in plant response to inoculation with mycorrhizal fungi.Ecology Letters13(3):394‑407. https://doi.org/10.1111/j.1461-0248.2009.01430.x
- Mycorrhizal Networks.Springerhttps://doi.org/10.1007/978-94-017-7395-9
- Impacts of species richness on productivity in a large-scale subtropical forest experiment.Science362(6410):80‑83. https://doi.org/10.1126/science.aat6405
- An Introduction to Population Biology.Yale Univ. Press
- Effect of diversity on growth, mortality, and loss of resilience to extreme climate events in a tropical planted forest experiment.Scientific Reports8(1):1‑10. https://doi.org/10.1038/s41598-018-33670-x
- High plant diversity is needed to maintain ecosystem services.Nature477(7363):199‑202. https://doi.org/10.1038/nature10282
- Biodiversity increases the resistance of ecosystem productivity to climate extremes.Nature526(7574):574‑577. https://doi.org/10.1038/nature15374
- Benefits of increasing plant diversity in sustainable agroecosystems.Journal of Ecology105(4):871‑879. https://doi.org/10.1111/1365-2745.12789
- Linking the influence and dependence of people on biodiversity across scales.Nature546(7656):65‑72. https://doi.org/10.1038/nature22899
- Phosphorus acquisition strategies within arbuscular mycorrhizal fungal community of a single field site.Plant and Soil276(1-2):163‑176. https://doi.org/10.1007/s11104-005-4274-0
- The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility.Biology and Fertility of Soils37(1):1‑16. https://doi.org/10.1007/s00374-002-0546-5
- Conspecific negative density dependence and forest diversity.Science336(6083):904‑907. https://doi.org/10.1126/science.1220269
- Functioning of mycorrhizal associations along the mutualism-parasitism continuum.The New Phytologist135(4):575‑585. https://doi.org/10.1046/j.1469-8137.1997.00729.x
- The continuum concept remains a useful framework for studying mycorrhizal functioning.Plant and Soil363(1):411‑419. https://doi.org/10.1007/s11104-012-1406-1
- Genotypic richness and dissimilarity opposingly affect ecosystem functioning.Ecology Letters14(6):537‑545. https://doi.org/10.1111/j.1461-0248.2011.01613.x
- TRY - a global database of plant traits.Global Change Biology17(9):2905‑2935. https://doi.org/10.1111/j.1365-2486.2011.02451.x
- Relationship Between Belowground Carbon Allocation and Nitrogen Uptake in Saplings Varies by Plant Mycorrhizal Type.Frontiers in Forests and Global Change2:81. https://doi.org/10.3389/ffgc.2019.00081
- Belowground carbon trade among tall trees in a temperate forest.Science352(6283):342‑344. https://doi.org/10.1126/science.aad6188
- The influence of arbuscular mycorrhizae on the relationship between plant diversity and productivity.Ecology Letters3(2):137‑141. https://doi.org/10.1046/j.1461-0248.2000.00131.x
- Functional complementarity in the arbuscular mycorrhizal symbiosis.New Phytologist147(2):233‑235. https://doi.org/10.1046/j.1469-8137.2000.00710.x
- Measurement of soil microbial biomass phosphorus by an anion exchange membrane method.Soil Biology and Biochemistry27(10):1353‑1357. https://doi.org/10.1016/0038-0717(95)00057-L
- Plant-soil feedbacks provide an additional explanation for diversity-productivity relationships.Proceedings of the Royal Society B: Biological Sciences279(1740):3020‑3026. https://doi.org/10.1098/rspb.2012.0285
- Plant diversity drives soil carbon storage by increased soil microbial activity.Nature Communications6:6707. https://doi.org/10.1038/ncomms7707
- Networks of power and influence: the role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning.Canadian Journal of Botany82(8):1016‑1045. https://doi.org/10.1139/b04-060
- Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats.Nature Communications6(1):1‑7. https://doi.org/10.1038/ncomms7936
- Spatial separation of litter decomposition and mycorrhizal nitrogen uptake in a boreal forest.New Phytologist173(3):611‑620. https://doi.org/10.1111/j.1469-8137.2006.01936.x
- Similar below‐ground carbon cycling dynamics but contrasting modes of nitrogen cycling between arbuscular mycorrhizal and ectomycorrhizal forests.New Phytologist213(3):1440‑1451. https://doi.org/10.1111/nph.14206
- An Ecological Loop: Host Microbiomes across Multitrophic Interactions.Trends in Ecology & Evolutionhttps://doi.org/10.1016/j.tree.2019.07.011
- Tree species richness increases ecosystem carbon storage in subtropical forests.Proceedings of the Royal Society B: Biological Sciences285(1885). https://doi.org/10.1098/rspb.2018.1240
- Partitioning selection and complementarity in biodiversity experiments.Nature412(6842):72‑76. https://doi.org/10.1038/35083573
- Biodiversity and ecosystem functioning: current knowledge and future challenges.Science294(5543):804‑808. https://doi.org/10.1126/science.1064088
- Soil microbes promote complementarity effects among co‐existing trees through soil nitrogen partitioning.Functional Ecology32(7):1879‑1889. https://doi.org/10.1111/1365-2435.13109
- Recommendations for establishing global collaborative networks in soil ecology.Soil Organisms91(3):73. https://doi.org/10.25674/so91iss3pp73
- Influence of phylogeny on fungal community assembly and ecosystem functioning.Science316(5832):1746‑1748. https://doi.org/10.1126/science.1143082
- Evolutionary history resolves global organization of root functional traits.Nature555(7694):94‑97. https://doi.org/10.1038/nature25783
- Measurement of phosphorus in the soil microbial biomass: a modified procedure for field soils.Soil Biology and Biochemistry18(4):437‑443. https://doi.org/10.1016/0038-0717(86)90050-7
- Biodiversity-multifunctionality relationships depend on identity and number of measured functions.Nature Ecology & Evolution2(1):44‑49. https://doi.org/10.1038/s41559-017-0391-4
- Root depth distribution and the diversity-productivity relationship in a long‐term grassland experiment.Ecology94(4):787‑793. https://doi.org/10.1890/12-1399.1
- Declining biodiversity can alter the performance of ecosystems.Nature368(6473):734‑737. https://doi.org/10.1038/368734a0
- Below-ground complementarity effects in a grassland biodiversity experiment are related to deep-rooting species.Journal of Ecology106(1):265‑277. https://doi.org/10.1111/1365-2745.12877
- A large and persistent carbon sink in the world’s forests.Science333(6045):988‑993. https://doi.org/10.1126/science.1201609
- A million and more trees for science.Nature Ecology & Evolution2(5):763‑766. https://doi.org/10.1038/s41559-018-0544-0
- Mycorrhizas: Anatomy and Cell Biology.NRC Research Presshttps://doi.org/10.1017/S0953756205213126
- Blurred boundaries: lifestyle lessons from ectomycorrhizal fungal genomes.Trends in Genetics27(1):14‑22. https://doi.org/10.1016/j.tig.2010.10.005
- Phylogenetic trait conservatism and the evolution of functional trade-offs in arbuscular mycorrhizal fungi.Proceedings of the Royal Society B: Biological Sciences276(1676):4237‑4245. https://doi.org/10.1098/rspb.2009.1015
- Modes of functional biodiversity control on tree productivity across the European continent.Global Ecology and Biogeography25(3):251‑262. https://doi.org/10.1111/geb.12406
- Biodiversity and ecosystem functioning relations in European forests depend on environmental context.Ecology Letters20(11):1414‑1426. https://doi.org/10.1111/ele.12849
- Mycorrhizas and nutrient cycling in ecosystems-a journey towards relevance?New Phytologist157(3):475‑492. https://doi.org/10.1046/j.1469-8137.2003.00704.x
- Impacts of biodiversity loss escalate through time as redundancy fades.Science336(6081):589‑592. https://doi.org/10.1126/science.1217909
- More closely related plants have more distinct mycorrhizal communities.AoB Plants6https://doi.org/10.1093/aobpla/plu051
- Uav-based high resolution thermal imaging for vegetation monitoring, and plant phenotyping using ici 8640 p, flir vue pro r 640, and thermomap cameras.Remote Sensing11(3):330. https://doi.org/10.3390/rs11030330
- Surrounding species diversity improves subtropical seedlings’ carbon dynamics.Ecology and Evolution8(14):7055‑7067. https://doi.org/10.1002/ece3.4225
- Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment.Nature468(7323):553‑556. https://doi.org/10.1038/nature09492
- The iDiv Ecotron-a flexible research platform for multitrophic biodiversity research.Ecology and Evolution11(21):15174‑15190. https://doi.org/10.1002/ece3.8198
- Persistence of soil organic matter as an ecosystem property.Nature478(7367):49‑56. https://doi.org/10.1038/nature10386
- Soil microbes drive the classic plant diversity-productivity pattern.Ecology92(2):296‑303. https://doi.org/10.1890/10-0773.1
- How accurately can soil organic carbon stocks and stock changes be quantified by soil inventories?Biogeosciences8(5):1193‑1212. https://doi.org/10.5194/bg-8-1193-2011
- Ecosystem Function of Biodiversity: A Summary.Springer Science & Business Mediahttps://doi.org/10.1007/978-3-642-58001-7_24
- Warming alters energetic structure and function but not resilience of soil food webs.Nature Climate Change7(12):895‑900. https://doi.org/10.1038/s41558-017-0002-z
- Mycorrhizal networks: des liaisons dangereuses?Trends in Ecology & Evolution21(11):621‑628. https://doi.org/10.1016/j.tree.2006.07.003
- Drought modulates interactions between arbuscular mycorrhizal fungal diversity and barley genotype diversity.Scientific Reports9(1). https://doi.org/10.1038/s41598-019-45702-1
- Net transfer of carbon between ectomycorrhizal tree species in the field.Nature388(6642):579‑582. https://doi.org/10.1038/41557
- Mycorrhizal networks: a review of their extent, function, and importance.Canadian Journal of Botany82(8):1140‑1165. https://doi.org/10.1139/b04-116
- Spatial differences in acquisition of soil phosphate between two arbuscular mycorrhizal fungi in symbiosis with Medicago truncatula.New Phytologist147(2):357‑366. https://doi.org/10.1046/j.1469-8137.2000.00695.x
- Mycorrhizal symbiosis.Academic Presshttps://doi.org/10.1016/B978-0-12-370526-6.X5001-6
- Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality.Nature536(7617):456‑459. https://doi.org/10.1038/nature19092
- Global mycorrhizal plant distribution linked to terrestrial carbon stocks.Nature Communications10(1). https://doi.org/10.1038/s41467-019-13019-2
- Climatic controls of decomposition drive the global biogeography of forest-tree symbioses.Nature569(7756):404‑408. https://doi.org/10.1038/s41586-019-1128-0
- Environmental drivers of ectomycorrhizal communities in Europe's temperate oak forests.Molecular Ecology23(22):5628‑5644. https://doi.org/10.1111/mec.12947
- Soil organic carbon pools in the northern circumpolar permafrost region.Global Biogeochemical Cycles23(2). https://doi.org/10.1029/2008GB003327
- Plant diversity drives soil microbial biomass carbon in grasslands irrespective of global environmental change factors.Global Change Biology21(11):4076‑4085. https://doi.org/10.1111/gcb.13011
- Reduced feeding activity of soil detritivores under warmer and drier conditions.Nature Climate Change8(1):75‑78. https://doi.org/10.1038/s41558-017-0032-6
- Advancing biodiversity-ecosystem functioning science using high-density tree-based experiments over functional diversity gradients.Oecologia174(3):609‑621. https://doi.org/10.1007/s00442-013-2815-4
- Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity.Nature396(6706):69‑72. https://doi.org/10.1038/23932
- Functional traits in mycorrhizal ecology: their use for predicting the impact of arbuscular mycorrhizal fungal communities on plant growth and ecosystem functioning.New Phytologist174(2):244‑250. https://doi.org/10.1111/j.1469-8137.2007.02041.x
- Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems.Journal of Ecology97(6):1139‑1150. https://doi.org/10.1111/j.1365-2745.2009.01570.x
- Mycorrhizal ecology and evolution: the past, the present, and the future.New Phytologist205(4):1406‑1423. https://doi.org/10.1111/nph.13288
- Jack-of-all-trades effects drive biodiversity-ecosystem multifunctionality relationships in European forests.Nature Communications7(1):1‑11. https://doi.org/10.1038/ncomms11109
- Contributions of a global network of tree diversity experiments to sustainable forest plantations.Ambio45(1):29‑41. https://doi.org/10.1007/s13280-015-0685-1
- Mycorrhizal fungal identity and richness determine the diversity and productivity of a tallgrass prairie system.New Phytologist172(3):554‑562. https://doi.org/10.1111/j.1469-8137.2006.01854.x
- Complementarity in both plant and mycorrhizal fungal communities are not necessarily increased by diversity in the other.Journal of Ecology103(5):1233‑1244. https://doi.org/10.1111/1365-2745.12452
- Plant species richness does not attenuate responses of soil microbial and nematode communities to a flood event.Soil Biology and Biochemistry89:135‑149. https://doi.org/10.1016/j.soilbio.2015.07.001
- Ecological linkages between aboveground and belowground biota.Science304(5677):1629‑1633. https://doi.org/10.1126/science.1094875
- Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions.Basic and Applied Ecology23:1‑73. https://doi.org/10.1016/j.baae.2017.06.002
- Spatial complementarity in tree crowns explains overyielding in species mixtures.Nature Ecology & Evolution1(4):1‑7. https://doi.org/10.1038/s41559-016-0063
- Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: results from long‐term field experiments.Ecology Letters12(5):452‑461. https://doi.org/10.1111/j.1461-0248.2009.01303.x
- The overlooked role of facilitation in biodiversity experiments.Trends in Ecology & Evolution32(5):383‑390. https://doi.org/10.1016/j.tree.2017.02.011
- Species richness promotes ecosystem carbon storage: evidence from biodiversity-ecosystem functioning experiments.Proceedings of the Royal Society B287(1939):20202063. https://doi.org/10.1098/rspb.2020.2063
- Aboveground litter inputs determine carbon storage across soil profiles: a meta-analysis.Plant and Soil1-16https://doi.org/10.1007/s11104-021-04881-5