|
Research Ideas and Outcomes :
Research Article
|
|
Corresponding author: Andrey Vyshedskiy (vysha@bu.edu)
Received: 29 Aug 2017 | Published: 31 Aug 2017
© 2017 Andrey Vyshedskiy, Shreyas Mahapatra, Rita Dunn
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Vyshedskiy A, Mahapatra S, Dunn R (2017) Linguistically deprived children: meta-analysis of published research underlines the importance of early syntactic language use for normal brain development. Research Ideas and Outcomes 3: e20696. https://doi.org/10.3897/rio.3.e20696
|
|
Abstract
We analyzed all published reports of individuals not exposed to syntactic language until puberty: two feral children, who grew up without hearing any language, and eight deaf linguistic isolates, who grew up communicating to their families using homesign or kitchensign, a system of gestures which allows them to communicate simple commands but lacks much in the way of syntax. A common observation in these individuals is the lifelong difficulty understanding syntax and spatial prepositions, even after many years of rehabilitation. This debilitating condition stands in stark contrast to linguistic isolates’ performance on memory as well as semantic tests: they could easily remember hundreds of newly learned words and identify previously seen objects by name. The lack of syntactic language comprehension in linguistic isolates may stem from inability to understand words and/or grammar or inability to mentally synthesize known objects into novel configurations. We have previously shown that purposeful construction of novel mental images is the function of the lateral prefrontal cortex (LPFC) ability to dynamically control posterior cortex neurons (Vyshedskiy et al. 2017). Here we have ranked all tests performed on linguistic isolates by their reliance on the LPFC control of the posterior cortex: a) the amount of posterior cortex territory that needs to be recruited by the LPFC and b) the number of disparate objects that have to be combined together by the LPFC in order to answer the test question. According to our analysis, linguistic isolates performed well in all tests that did not involve the LPFC control of the posterior cortex, showed decreasing scores in tests that involved greater recruitment of the posterior cortex by the LPFC, and failed in tests that involved greatest recruitment of posterior cortex necessary for mental synthesis of multiple objects. This pattern is consistent with inadequate frontoposterior connections in linguistic isolates. We discuss implications of these findings for the importance of early syntactic language exposure in formation of frontoposterior connections.
Keywords
Language acquisition, critical period, language acquisition critical period, linguistically deprived, linguistic isolates, first language, first language acquisition, Genie, feral children, linguistic isolates.
Introduction
There is general consensus that early time-sensitive exposure to syntactic language is important for acquisition of the full extent of a complex syntactic language (
The best studied case of a completely linguistically isolated child is that of Genie (real name: Susan M. Wiley) who was isolated starting at 20 months of age until she was rescued at the age of 13 years 7 months. She was locked inside a bedroom in Los Angeles, strapped to a child’s toilet during the day and bound inside a crib with her arms and legs immobilized on most nights (
Genie’s case has been extensively compared to that of Victor of Aveyron, an eighteenth-century French child whose life similarly became a case study for research in delayed first-language acquisition. Victor of Aveyron was a feral child who apparently lived much of his childhood alone in the woods before being found wandering the woods near Saint-Sernin-sur-Rance in France. His case was taken up by a young physician, Jean Marc Gaspard Itard, who worked with the boy for five years. Itard named the wild child Victor, and estimated that the boy was 13 years old when he initiated the study. Itard, who was interested in determining what Victor could learn, devised procedures to teach the boy words and recorded his progress (
Syntactic isolation of children stems from various causes and may occur even in a loving and cohesive family. About 90% of all congenitally deaf children are born to hearing parents (
Deaf children who grow up using homesign for communication must be distinguished from deaf children developing in a community of other deaf children, as they are known to be able to independently invent a syntactic sign language of their own. In 1980, following the Sandinista revolution, the Nicaraguan government opened several vocational schools for deaf children. By 1983 there were over 400 students in the two schools. The school program emphasized spoken Spanish and lip reading, and discouraged the use of signs by teachers. The program failed and students were unable to learn the Spanish language in such a manner. However, the school provided fertile ground for deaf students to communicate with each other. In this process, children gradually spontaneously generated a new sign language, complete with syntax, verb agreement and other conventions of grammar (
Syntactically isolated children may also include those who are exposed to syntactic language but do not use it either for external or for internal communications. For the most part this group is formed by children with ASD. The primary problem in these children may be neurological, leading to a secondary problem of significant language delay. In turn, lack of syntactic language use during the critical period may result in the tertiary cognitive problems. About two thirds of children with ASD eventually demonstrate significant cognitive impairment (
Additional evidence for the importance of early language exposure for normal cognitive development comes from the controlled randomized study of the orphaned Romanian children (
In this manuscript we initially focus on the most definitive cases of language isolation: children who have been linguistically isolated until puberty and then subjected to rehabilitation attempts for several years in their adolescence or adulthood. These cases of feral children and deaf linguistic isolates present the best opportunity to delineate the effect of syntactic language deprivation on the developing brain. In the discussion, we compare our findings to other children with relatively shorter first syntactic language deprivation periods.
Material and methods
We performed PubMed and GoogleScholar search in May of 2017 for all reports of linguistic isolates. The search yielded reports of ten individuals not exposed to a syntactic language until puberty, which were subsequently tested as adults following several years of focused rehabilitation: two reports of feral children and eight reports of deaf linguistic isolates. Performance of these individuals in all kinds of linguistic and nonverbal tests is reviewed in this manuscript in terms of their PFC ability to manipulate and control their imaginary visual experience.
Assessment of linguistic test questions
A panel of three neuroscientists classified all language comprehension and nonverbal tasks administered to linguistic isolates according to the minimal neurobiological requirements necessary to obtain the correct answer (
(2.1) Establishment of the type of object modification: For any object in the mind’s eye, we can voluntarily change its color, size, position in space or rotation. The mechanisms of these processes involve PFC-controlled modification of the object’s representation in the posterior cortex. As related to the posterior cortex neuronal territory, such mechanisms can be classified into three classes: 1) those that do not involve any object modification; 2) those that involve modification of activity in only the ventral visual cortex (Fig.
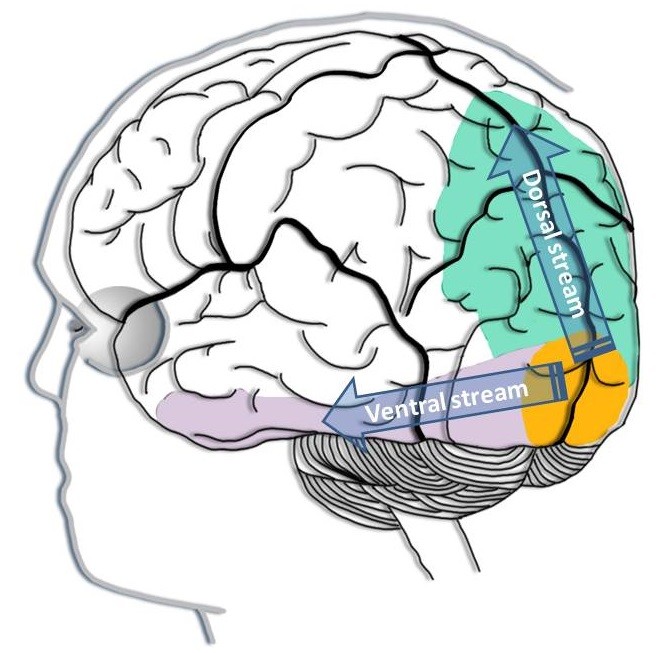
Visual information processing in the cortex. From the primary visual cortex (V1, shown in yellow), the visual information is passed in two streams. The neurons along the ventral stream also known as the ventral visual cortex (shown in purple) are primarily concerned with what the object is. The ventral visual stream runs into the inferior temporal lobe. The neurons along the dorsal stream also known the dorsal visual cortex (shown in green) are primarily concerned with where the object is. The dorsal visual stream runs into the parietal lobe.
Language comprehension tests
1. Find the named object. A common task requires the subject to find a named object from a variety of physical objects (or pictures) to choose from. These tasks are based of subject’s ability to compare the internal representation of the target object with all the possible solution objects, primarily relying on matching from memory. Since this type of question does not require any mental combination of disparate objects, it is assigned a NOB score of one. Furthermore, since this type of question does not require any modification of the object, it corresponds to the PCT score of zero (i.e. requiring the least amount of the posterior cortex territory).
2. Integration of modifiers in a single object. Another common comprehension test requires the subject to integrate a noun and an adjective. A subject may be asked to point out the picture with a {yellow/red/green} + circle placed among several decoy images with {yellow/red/green} + {triangle/square} thus forcing the integration of color and noun. Similarly, to integrate size and noun one may be asked to point to a {big/small} + {circle/triangle/square}. Neurologically, the integration of modifiers involves the modification of neurons encoding a single object and therefore has a NOB score of one. As color and/or size were modified, these tasks were given a PCT score of one. (modification is limited to the ventral visual cortex, see
2.1. Comparative. In a comparative test the subject may be shown two circles of different sizes and asked to point out the circle that is {bigger/smaller}. In terms of visual representation in the posterior cortex, the circles can be considered one at a time, making the comparative task a special case of integration of the size modifier task. Therefore, minimally, the comparative task involves the modification of neurons encoding a single object (the circle) and has a NOB score of one. In terms of the PCT, these questions were assigned a score of one (modification is limited to the category of size represented in the ventral visual cortex, see
2.2. Superlative. In a superlative test, the subjects may be shown several circles of different sizes not aligned by size and asked to point out the biggest/smallest circle. Similar to comparative task, superlative task is a special case of integration of the size modifier, it involves the modification of neurons encoding a single object and therefore has a NOB score of one. In terms of the PCT, these questions were assigned a score of one (modification is limited to the ventral visual cortex, see
3. Integration of number modifier. To integrate the number and noun the subject may be asked to point out a card with {two/three/four} circles among distractor cards with {two/three/four} + {squares/triangles}. Similar to comparative task, the integration of number modifier minimally involves the modification of neurons encoding the number: independently of whether the number is two or a million, only a single object (the circle) is visualized and therefore this test has a NOB score of one. Reports indicate that numerical information is represented by regions of the posterior parietal lobes (
3.1. Singular vs. plural. Singular vs. plural tasks involve showing several pictures and asking the subject to point out the correct picture: ‘Show me the flower’ versus ‘Show me the flowers. Neurologically, in terms of visual representation of object in the posterior cortex, these tasks are the special case of the integration of number modifier; they involve the modification of neurons encoding a single object (NOB score of one) and have a PCT score of two.
3.2. Negative vs. affirmative statements. To test understanding of simple negation the subject is shown a pair of pictures identical except for the presence or absence of some element. For example, the subject may be asked to indicate a picture in which the rabbit has a carrot vs. the one in which the rabbit does not have a carrot. Neurologically, in terms of visual representation of object in the posterior cortex, these tasks involve the modification of neurons encoding a single object (e.g., a carrot) and therefore have a NOB score of one. These questions are a special case of integration of number modifier, with the number set to zero. Accordingly, they were assigned a PCT score of two.
4. Mental rotation and modification of a single object’s location in space. A number of verbal and nonverbal tasks given to linguistic isolates require mental rotation or other modification of an object’s location in space. An example of a nonverbal mental rotation task is presented in Fig.
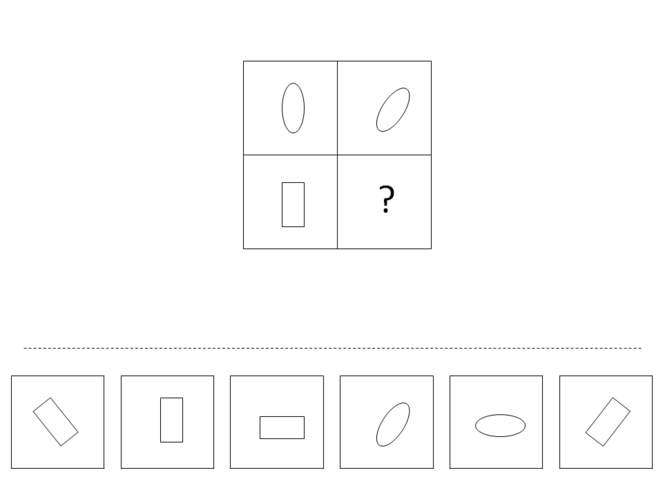
A typical question testing subject’s ability to mentally rotate an object is shown here as a 2x2 matrix with six answer choices displayed below the problem. The top row of the matrix indicates the rule: “the object in the right column is the result of 45° clockwise rotation.” Applying this rule to the bottom row, we arrive at the correct answer depicted on the right.
4.1 Copying stick and block structures. Copying stick and block structures test is a nonverbal test that relies on mental rotation of sticks or block pieces in order to align them with the instructions. Since mental manipulations involve one piece at a time, these tasks are limited to a single object and, consequently, assigned a NOB score of one. Mental rotation involve the PFC-coordinated activity in both the ventral and the dorsal visual cortices (
5. Mental synthesis of several objects.
5.1 Spatial prepositions. Understanding of spatial prepositions such as in, on, under, over, beside, in front of, behind requires a subject to superimpose several objects. For example, the request “to put a green box {inside/behind/on top of} the blue box” requires an initial mental simulation of the scene, only after which is it possible to correctly arrange the physical objects. An inability to produce a novel mental image of the green box {inside/behind/on top of} the blue box would lead to the use of trial-and-error, which in majority of cases will result in an incorrect arrangement. Understanding an instruction to “put the bowl {behind/in front of/on/under} the cup requires the combination of two objects and therefore is assigned a NOB score of two. Understanding an instruction to “put the cup in the bowl and on the table” requires the combination of three objects and therefore is assigned a NOB score of three. In general, a NOB score in mental synthesis questions is defined as the number of disparate objects that have to be imagined together.
When two disparate objects are imagined together their spatial relationship has to be encoded. Encoding of spatial information is the function of the dorsal visual cortex (
5.2 Flexible syntax. The order of words, or syntax (from Greek syn, meaning together, and taxis, meaning an ordering), is an essential quality of all human languages. A change in the word-order often completely changes the meaning of a sentence. For example, the phrase “a cat ate a mouse” and the phrase “a mouse ate a cat,” have very different connotations only because the word order was changed. To understand the exact meaning of these two sentences, the subject must mentally synthesize the objects of “a cat” and “a mouse.”
Flexible syntax is distinguished from rigid syntax, whereby the meaning of a word is always dictated by its position in a sentence. For example, if we all agree that the first noun in the “noun1 ate noun2” sentence will always be the eater and the second noun will always be the prey, then the meaning of the sentence can be inferred without mental synthesis of multiple objects. When you hear “a cat ate a mouse,” you automatically know that noun1 (a cat) was the eater and noun2 (a mouse) was the prey. You do not need to synthesize the image of a cat and a mouse together; you can simply infer the meaning from the sequence of words in the sentence. Of course this rule constricts the conversation to a single sentence structure: “noun1 ate noun2.” If you follow this structure strictly, you will always misinterpret sentences like “a cat was eaten by a mouse,” “a cat will be eaten by a mouse,” “this cat never eats mice,” since you will always think that noun1 (the cat) is the eater. Human languages are not bound by limitations of rigid syntax: both the first and the second nouns can be the ‘eaters’ (“a cat ate a mouse” versus “a cat was eaten by a mouse”). All human languages use flexible syntax and rely on mental synthesis of multiple objects to decode the meaning of a sentence.
Similar to spatial prepositions tasks, a NOB score in flexible syntax tasks is defined as the number of disparate objects that have to be imagined together and PCT score of two is assigned.
5.3 Before/after word order. To test understanding of before/after word order, subjects can be asked ‘to touch your nose after you touch your head.’ Similar to flexible syntax, this task relies on mental synthesis to simulate the process of touching the nose and the head. These tasks are given a NOB score of two and PCT score of two.
5.4 Active vs. passive. Subjects can be asked to point to a picture showing ‘The boy is pulling the girl’ vs. ‘The girl is pulled by the boy.’ Similar to flexible syntax, this task relies on mental synthesis to combine the boy and the girl in the mind’s eye. It is given a NOB score of two and PCT score of two.
5.5 Exact arithmetic abilities. Mentally finding the solution to a complex arithmetic problem such as mental two-digit addition (e.g. 43+56) and multiplication (e.g. 32x24) also relies on mental synthesis. This process is different from multiplication of single digit numbers (e.g. 2x4) that can be retrieving from memorized multiplication table (
6. Special cases
6.1. Classification tasks. Classification tasks involve sorting objects on the principles of gender, animacy, etc. Classification tasks involve no object combination and therefore were assigned a NOB score of one. The tasks also involve no modification of objects and therefore were assigned the PCT score of zero. Classification may be more challenging than similarly scored “Find the object” tasks, since the object’s class has to be determined, but this is not reflected in the scoring system used in this report.
6.2 Picture arrangement into a story. A typical test consists of three to six pictures with cartoon characters that can be arranged into a story. For example, picture A shows a boy running, picture B shows the boy falling, and picture C shows the boy crying. The pictures are presented to the subject in scrambled order with instructions to rearrange the pictures to make the best sense. A typical adult might approach this task by mentally animating the characters and generating a story using mental synthesis of multiple objects. However, in order to identify the minimal neurobiological requirements for the correct answer, we considered the following alternative. Most picture arrangement tasks rely on subject’s understanding of causality between events shown in the pictures. The inference of causality can be made from a memory of similar events watched on TV or a personal experience, e.g. running, falling and crying thereafter. The process of inferring the causality does not rely on combination or modification of objects and therefore was assigned a NOB score of one and the PCT score of zero.
The summary of NOB and PCT scores for all linguistic tests is provided in Table
The hierarchical classification of linguistic tests question by a NOB score and the PCT score.
|
Linguistic test |
Number of objects score |
Posterior cortex territory score |
|
Find the object |
1 |
0 |
|
Classification tasks |
1 |
0 |
|
Picture arrangement into a story |
1 |
0 |
|
Integration of size and color modifiers |
1 |
1 |
|
Comparative: e.g. ‘Which button is smaller?’ |
1 |
1 |
|
Superlative: e.g. ‘Which button is smallest out of five?’ |
1 |
1 |
|
Integration of number modifier |
1 |
2 |
|
Singular vs. plural |
1 |
2 |
|
Understanding of simple negation |
1 |
2 |
|
Mental rotation and modification of a single object’s location in space |
1 |
2 |
|
Copying stick and block structures |
1 |
2 |
|
Mental synthesis of multiple objects: Understanding of spatial prepositions, such as in, on, under, over, beside, in front of, behind, etc. |
2 |
2 |
|
Understanding of flexible syntax |
2 |
2 |
|
Before/after word order: |
2 |
2 |
|
Active vs. passive |
2 |
2 |
|
Exact arithmetic abilities: mental two-digit addition / multiplication |
2+ |
2 |
Comments to Figure 3 Linguistic isolates performance in verbal and nonverbal tests
Fig.
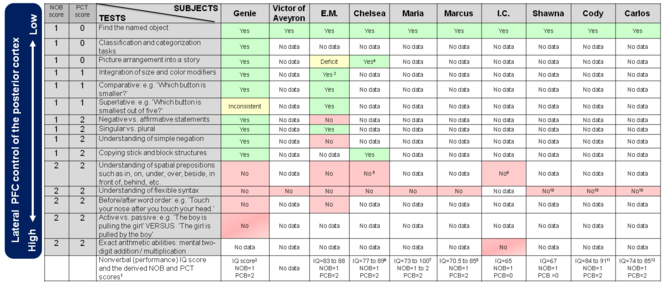
Comments:
1. The methodology for deriving a NOB and PCT scores from the performance IQ score was described (
2. Genie was tested with Raven’s Matrices and Wechsler Intelligence Scale for Children (WISC).
Using this limited information we attempted to map Genie’s IQ into a NOB and PCT scores:
A. WISC uses three sections to determine the performance score: Matrix Reasoning, Figure Weights, and Visual Puzzles. A person who can only answer test questions with a NOB score of one and PCT score of zero (i.e. “Find the same object” and “Amodal completion”) would receive scaled section scores of 2, 5, 3 respectively (
B. From
C. The latest nonverbal test was administered 5 years into rehabilitation “Genie was given the Coloured Progressive Matrices Test ... Genie’s overall score was 29.” (
Thus, in all three nonverbal tests that Genie was administered up to five years into rehabilitation Genie has invariably failed to demonstrate capacity for mental synthesis of multiple objects.
3. Grimshaw reports that E.M.’s “performance on both simple and complex modification tasks was above chance but imperfect from the earliest assessment. E.M. made more errors in complex modification (two adjectives) than in simple modification (one adjective). Almost all errors involved a failure to identify the correct shape—the modifiers were always correct” (
4. Curtiss reports that “She didn't perform as well as Genie on such tasks, but she could put picture sequences into a logical sequence” (personal communications).
5. Curtiss reports that “She did not appear to comprehend (or use) such elements” (personal communications).
6.
7.
8.
9. Subject I.C. was unable to use spatial signs (left/right, on top of/underneath) to describe the relationship between the objects.
10. According to
11. According to
12. According to
Results
Fig.
Genie was also tested on standardized nonverbal tests Raven’s Matrices (
Victor of Aveyron, an eighteenth-century French feral child entered his rehabilitation at the approximate age of 13 (Fig.
Studies of feral children, however, are highly problematic: isolation and childhood abuse can result in all sorts of psychological problems, which may confound conclusions drawn about linguistic abilities. Studies of congenitally deaf people, who had linguistic but not social deprivation, have fewer methodological weaknesses. To date, there have been only eight carefully documented cases of deaf subjects who were linguistically deprived until puberty.
While E.M.’s performance on simple linguistic tests that do not rely on mental synthesis was reasonably good, his performance on more complex tests that rely on mental synthesis of multiple objects was poor. Similar to Genie, E.M. had significant difficulty with spatial prepositions, even though his performance on this task was better than chance.
Another report was of Chelsea, a deaf woman isolated from syntactic linguistic input until she was 32, when her first exposure to spoken English was provided by successful auditory amplification (Fig.
Maria was administered Raven’s Standard Progressive Matrices test after three months and after 32 months exposure to ASL. Her score after three months’ exposure to ASL was below the fifth percentile for her age group, corresponding to a NOB score of one and the PCB score of two. After 32 months’ exposure to ASL, her performance increased to the fiftieth percentile for her age group (
Discussion
In this manuscript we analyzed all the published reports of linguistic isolates – individuals not exposed to syntactic language until puberty. The stark contrast between their lifelong difficulty in understanding spatial prepositions and syntax and their ability to learn hundreds of new words begs the question: is lack of syntactic language comprehension a language problem or an imagination problem? If linguistic isolates were not able to purposefully synthesize disparate objects into novel configurations in their mind’s eye, that could parsimoniously explain their lack of understanding flexible syntax, low nonverbal IQ, and their lifelong difficulty of processing spatial information. To test this hypothesis, we noticed that mental synthesis of disparate objects is the function of the lateral prefrontal cortex (LPFC) ability to control the posterior cortex neurons (
All of these observations are consistent with the hypothesis that understanding complex syntactic language is intimately connected to the control of the LPFC over objects encoded in the brain.
Objects as units of perception
Our visual world consists of meaningful, unified, and stable objects that move coherently as one piece. Objects, therefore, constitute the functional units of perception (
While the term “neuronal ensemble” is often used in a broad sense to refer to any population of neurons involved in a particular neural computation, in this discussion we will use the term more narrowly to mean a stable group of neurons which are connected by enhanced synapses and which encode specific objects. Thus, neuronal ensembles are internal equivalents of objects: while the latter are physical or external units of perception, the former are the internal units of perception; each object that has been seen and remembered by a subject is encoded by a neuronal ensemble.
The neurons of an ensemble are distributed throughout the posterior cortex (occipital, temporal, and parietal lobes). Most neurons encoding a stationary object are located within the ventral visual cortex (also known as the "what” stream) (
The role of the PFC
The PFC consists of two functionally distinct divisions. The ventromedial PFC is predominantly involved in emotional and social functions such as the control of impulse, mood, and empathy (
Lateral PFC-driven synchronization of neurons as the likely mechanism of internally-driven novel sensory experiences
We have previously hypothesized that the mechanism behind the mental synthesis of independent objects involves the LPFC acting in the temporal domain to synchronize the neuronal ensembles which encode those objects (
We have also hypothesized that the neurological process of modifier integration, whereby the LPFC modifies the activity of neurons in a single neuronal ensemble resulting in changes of an object’s perceived color or size, involves the LPFC acting in the temporal domain to synchronize the color encoding neurons in the visual area V4 with the object’s neuronal ensemble in the ventral visual cortex (
Synchronous frontoposterior connections
The Mental Synthesis theory also predicts that to synchronize neuronal ensembles dispersed throughout the posterior cortex, the LPFC must rely on synchronous connections to posterior cortex neurons (
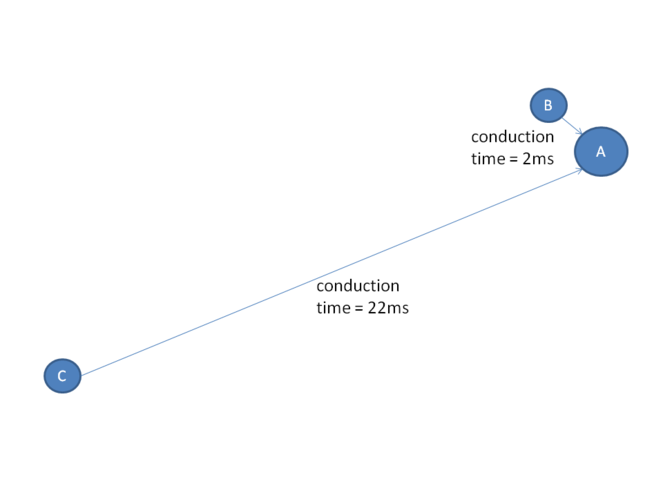
Synchronicity has to be understood in terms of synchronicity of the arrival of action potentials to a target neuron rather than absolute equality of action potential conduction times over different paths. Consider the following example: suppose neuron A is receiving excitatory input from neurons B and C via two different pathways (neuron A is the target neuron for both neurons B and C). Suppose that the action potential conduction time is 2ms from neuron B to neuron A and 22ms from neuron C to neuron A (i.e., the axonal pathway B-A has a significantly shorter conduction time than the axonal pathway C-A). Does it mean that the connections B-A and C-A are always asynchronous? No. The answer depends on the predominant neural activity rhythm in this network. At the firing rate of 50Hz (inter-spike interval of 20ms that correspond to Gamma rhythm), neurons B and C can actually be considered synchronous in relationship to neuron A: consider a train of action potentials synchronously fired by neurons B and C. The first action potential from neuron B will reach neuron A in 2ms and the first action potential from neuron C will reach neuron A in 22ms. Obviously, there would be no coincidence in the arrival times of the 1st action potentials from neurons B and C. However the second action potential from neuron B will arrive to neuron A in 22ms, concurrently with the 1st action potential from neuron C. Thus, starting with the second action potential, neuron A will receive synchronous activation from neurons B and C. The synchronous activation has a significantly greater probability of enhancing synaptic connections between neurons A and B, and A and C (Hebbian learning: ‘neurons that fire together, wire together’ (
Ontogenetic acquisition of synchronous frontoposterior connections
Myelination is nearly completed by birth in most species in which the young are relatively mature and mobile from the moment of birth, such as wild mice (
There is significant evidence that neural activity can affect myelination. The number of myelin-forming cells in the visual cortex of rats increases by 30% when the rats are raised in an environment that is enriched by additional play objects and social interaction (
Environmental effects on axonal myelination extend beyond animal studies. In human infants, early experiences increase myelination in the frontal lobes in parallel with improved performance on cognitive tests (
To sum up, synchronicity in at least some neuronal networks seems to be achieved via differential myelination and myelination may be experience-dependent. In fact, considering the many variables affecting conduction delays in an adult brain and the hundreds of millions of frontoposterior connections, genetic instruction alone would seem inadequate to specify the optimal conduction velocity in every axon. Accordingly, the development of synchronous frontoposterior connections likely involves adjustment of the conduction velocity in individual fibers via an experience-dependent process.
The experience-dependent developmental mechanisms have been clearly demonstrated for the visual, auditory, somatosensory and motor systems in virtually all species, from humans to Drosophila (
A common thread between these experience-dependent processes is that the ontogenetic experience essential for the formation of the neurological network is the same experience for which this network is naturally used in adulthood: stimulation of eyes by light is essential for formation of visual cortical connections, auditory stimulation is essential for formation of auditory cortex connections, motor practice is essential for formation of cortical motor connections, etc. Thus, it is compelling to hypothesize that cortical connections, which are essential for mental synthesis of multiple objects in adulthood are acquired as a result of the childhood mental synthesis exercises. Noting that conscious purposeful mental synthesis of disparate objects is primarily stimulated by syntactic language – whether it is internal language when we are thinking to ourselves or external language when we are involved in conversation – we would predict that the synchronous frontoposterior connections can only be acquired as a result of childhood exposure to syntactic language. The lack of syntactic language during the sensitive period of the PFC plasticity would result in asynchronous frontoposterior connections and mental synthesis disability.
In fact, this is exactly what we have observed in all linguistic isolates. Their lifelong mental synthesis disability is consistent with the critical role of early syntactic language use for establishment of synchronous frontoposterior connections. Childhood use of syntactic language (both externally and internally) likely provides the necessary training mechanism for development of synchronous connections between the LPFC and the sensory memory stored as neuronal ensembles in the posterior cortex. Without those synchronous connections having developed before the closure of the period of plasticity, the LPFC executive remains forever disabled at performing precise phase control of neurons, which are located over the large parts of the posterior cortex. The millions of strings connecting the LPFC puppeteer to its puppets (the neuronal ensembles in the posterior cortex) remain ill-adjusted, with the LPFC unable to control synchronization of independent neuronal ensembles and, therefore, unable to synthesize novel mental images. A child involved in a normal syntactic conversation (either with another person or with himself) internally manipulates mental images in his/her mind and during this process adjusts conduction velocity to achieve synchronicity in the neural networks connecting the LPFC to widespread regions of the posterior cortex.
The critical role of the syntactic language in formation of synchronous frontoposterior connections and mental synthesis is consistent with findings in late first-language learners and nonverbal individuals with autism. Elissa Newport and Rachel Mayberry studied the acquisition of sign language in deaf individuals differing in age of exposure: some were exposed to sign language from birth, while other children first learned sign language at school. These studies were conducted with adults who have been using sign language for at least twenty years. The results of the studies consistently indicated a negative correlation between the age of sign language acquisition and ultimate proficiency: those exposed to sign language from birth performed best, and late learners — worst, on all production and comprehension tests (
Prelingual deafness is such a serious concern that the US government has enacted laws to identify affected newborns. In 1999, the US congress passed the “Newborn and Infant Hearing Screening and Intervention Act,” which gives grants to help states create hearing screening programs for newborns. Otoacoustic Emissions Testing is usually done at birth, followed by an Auditory Brain Stem Response if the Otoacoustic Emissions test results indicated possible hearing loss. Such screening allows parents to expose deaf children to a formal sign language as early as possible and therefore avoid any delay in introduction to syntactic language.
Problems observed in late first-language learners also afflict many individuals with autism, leading to what is commonly described as “stimulus overselectivity,” “tunnel vision,” or “lack of multi-cue responsivity” (
The duration of the critical period for acquisition of synchronous frontoposterior connections
The period of plasticity for the development of synchronous frontoposterior connections seems to close in several stages. Based on the randomized controlled study of institutionalized Romanian children, the plasticity seems to already decline before the age of two: children placed in foster care and therefore exposed to complex syntactic language before the age of 2 showed increased myelination and performed better in mental integration tasks compared to children who have been placed in foster care after the age of 2 (
The next significant decline in plasticity is around the age of five. When the left hemisphere is surgically removed before the age of five (to treat cancer or epilepsy), patients often attain normal cognitive functions in adulthood (using the one remaining hemisphere). Removal of the left hemisphere after the age of five often results in significant impairment of syntactic language and tasks requiring mental synthesis (
Further reduction of plasticity occurs by the time of puberty; a lack of experience in syntactic speech before puberty likely results in a consolidation of widely desynchronized frontoposterior networks and prevents individuals from subsequent acquisition of mental synthesis. While parts of the PFC network retain some plasticity for a significantly longer period of time, since myelination of the PFC continues into the third decade of life or later (
Is there a neurological dissociation between language and cognition?
It is common among linguists to discuss the “language module” in the brain. In support of the existence of the “language module,” linguists enlist patients with apparent dissociation between language and cognition (
The word “language” has highly divergent meanings in different contexts and disciplines. In informal usage, a language is usually understood as a culturally specific communication system (German, French, or Spanish Language), i.e. understanding of individual words without any reference to understanding of flexible syntax. On the other hand, in linguistics, the term “language” is used quite differently to encompass both understanding of words and flexible syntax. Fitch, Hauser, & Chomsky (
Mental synthesis mediated by the LPFC is the best candidate for the “abstract core of computational operations, central to language and probably unique to humans” described by Fitch, Hauser, & Chomsky (
All human languages are recursive: they allow high fidelity transmission of infinite number of novel images with the use of a finite number of words, Fig.
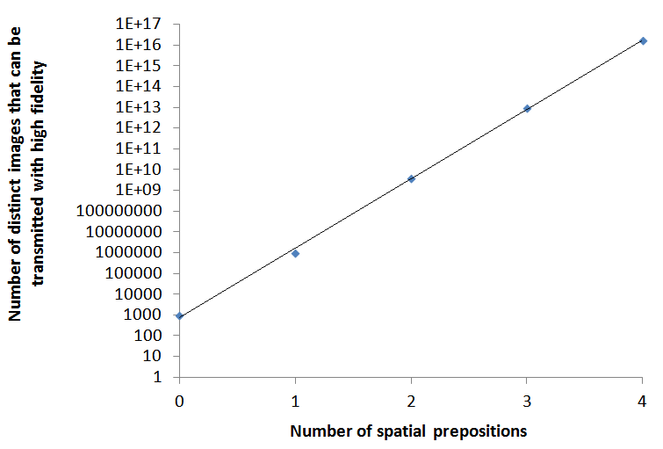
Flexible syntax, prepositions, adjectives, verb tenses, and other common elements of grammar, all facilitate the human ability to communicate an infinite number of novel images with the use of a finite number of words. The graph shows the number of distinct images that can be transmitted with high fidelity in a communication system with 1,000 nouns as a function of the number of spatial prepositions. In a communication system with no spatial prepositions and other recursive elements, 1000 nouns can communicate 1000 images to a listener. Adding just one spatial preposition allows for the formation of three-word phrases (such as: ‘a bowl behind a cup’ or ‘a cup behind a bowl’) and increases the number of distinct images that can be communicated to a listener from 1000 to one million (1000x1x1000). Adding a second spatial preposition and allowing for five-word sentences of the form object-preposition-object-preposition-object (such as: a bowl on a cup behind a plate) increases the number of distinct images that can be communicated to four billion (1000x2x1000x2x1000). The addition of a third spatial preposition increases the number of distinct images to 27 trillion (1000x3x1000x3x1000x3x1000), and so on. In general, the number of distinct images communicated by three-word sentences of the structure object-preposition-object equals the number of object-words times the number of prepositions times the number of object-words. A typical language with 1000 nouns and 100 spatial prepositions can theoretically communicate 1000101 x 100100 distinct images. This number is significantly greater than the total number of atoms in the universe. For all practical purposes, an infinite number of distinct images can be communicated by a syntactic communication system with just 1000 words and a few prepositions. Prepositions, adjectives, and verb tenses dramatically facilitate the capacity of a syntactic communication system with a finite number of words to communicate an infinite number of distinct images. Linguists refer to this property of human languages as recursion. The “infiniteness” of human language has been explicitly recognized by “Galileo, Descartes, and the 17th-century ‘philosophical grammarians’ and their successors, notably von Humboldt” (
1) Some expressive language is completely automated, for example, coprolalia sometimes observed in patients with Tourette syndrome involves spontaneous and articulate uttering of expletives in an uncontrollable manner (
2) Tape recorder language and reading poetry by heart can sometimes be completely automated, prerecorded through repetition (
3) Echolalia in its profound form is automatic and effortless, imitative behavior whereby sounds or actions are imitated without explicit awareness (
4) Confabulations involve the production of fabricated memories without the conscious intention to deceive. A liar would commonly initially simulate a story by using LPFC-controlled mental synthesis of multiple objects and check it for consistency before delivering the story to the listeners. The confabulator, by definition, does not have a pre-simulated story that could indicate the ability of the LPFC to ascertain control over the posterior cortex. Confabulatory tendencies are often interpreted as an indicator of a disconnection between PFC and the posterior cortex (
5) Many people report the experience of reading aloud a book to a child and thinking of something else. In this case their LPFC is likely driving their mental synthesis while the book’s sensory input is driving the Broca’s area speech production and the right hemisphere's prosody in the bottom-up manner.
6) The vivid, original, bizarre, emotional, and story-like dreams that most people remember when they wake up occur during rapid eye movement (REM) sleep. These dreams consist of existing fragments of memory put into various new combinations and are often erroneously assumed to be created by the LPFC via the mechanism similar to mental synthesis. For example, Darwin associates imagination with dreaming: “The imagination is one of the highest prerogatives of man. By this faculty he unites former images and ideas, independently of the will, and thus creates brilliant and novel results... Dreaming gives us the best notion of this power... The value of the products of our imagination depends of course on the number, accuracy, and clearness of our impressions, on our judgment and taste in selecting or rejecting the involuntary combinations, and to a certain extent on our power of voluntarily combining them. As dogs, cats, horses, and probably all the higher animals, even birds have vivid dreams, and this is shown by their movements and the sounds uttered, we must admit that they possess some power of imagination.” (
Two lines of evidence point toward the mechanism of dreams that does not depend on the LPFC. First, neuroimaging observations indicate that the LPFC is inactive during sleep, both slow-wave sleep and REM sleep (
We conclude that it is often impossible for an external observer to reliably infer internal imagery of the speaker based on expressive language. The speech may be automatic and independent of internal imagery, triggered spontaneously or driven by sensory input. Even when words reflect internal images, they may not inform of the LPFC control over imagination since even the novel images may have formed spontaneously as in dreaming or hallucination. There is no accurate method for external assessment of mental synthesis with respect to expressive language. While understanding of flexible syntax commonly requires LPFC-controlled mental synthesis, expressive language does not.
Several other bottom-up processes are often erroneously assumed to be indicative of the LPFC-controlled mental synthesis:
1) Object recognition / perceptual closure. Recognition of pictures with incomplete visual information is often erroneously assumed to be under the control of the LPFC. While there is no doubt that objects encoded in the posterior cortex can be primed by a category encoded in the PFC, there is strong evidence of bottom-up-driven PFC-independent process in perceptual closure tests, such as Moony faces test (
2) Drawing interpretation. The tendency to perceive objects from lines and colors has been used extensively in psychological tests such as blot test. It is often as easy to recognize an object from meaningless colors or scribbles of lines as it is from a cloud in the sky. Furthermore, once we recognize an object in a drawing, we tend to presume knowing the inclinations of the artist’s mind. However, the artist may have produced the drawing automatically or just spilled some colors as in blot test pictures.
Even some representational drawings can be fully automatic. For example, instrumental training (a variation of Pavlovian training) has been used to train elephants to produce representational works of art. Patients with visual agnosia can copy a drawing without any awareness of the type of object in the picture. The famous patient known in literature as pilot John faithfully copied a representation of St. Paul Cathedral without knowing the object in the picture (
Conclusions
It is commonly believed that there is a strong association between early language acquisition and normal cognitive development (critical period hypothesis) (
As is the case with ontogenetic development of many other neurological systems from muscle innervation to the development of sensory systems, nature’s intent must be complemented by adequate nurture. What is highly unusual about the ontogenetic acquisition of mental synthesis is that the necessary experience is provided by the exposure to a purely cultural phenomenon: a syntactic language. For the normal development of vision, light reflected from surrounding objects has to reach the retina, but that occurs whenever it is light, independent of cultural exposure; for the normal development of the muscular system, the trophic factors released by muscles have to reach their neurons, but that occurs whenever a child is moving – the stimulation to neurons comes naturally even when a child is growing alone in a forest (
Acknowledgements
We wish to thank Dr. Petr Ilyinskii for productive discussion and scrupulous editing of this manuscript and Dr. Irene Piryatinsky for help with interpretation of IQ tests.
Funding program
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
- Early experience alters brain function and structure.Pediatrics113(4):846‑857. https://doi.org/10.1542/peds.113.4.846
- Proliferation of oligodendrocyte precursor cells depends on electrical activity in axons.Nature361(6409):258‑258. https://doi.org/10.1038/361258a0
- Experience-dependent developmental plasticity in the optic lobe of Drosophila melanogaster.Journal of Neuroscience17(4):1493‑1504.
- Age-related changes in frontal and temporal lobe volumes in men: a magnetic resonance imaging study.Archives of General Psychiatry58(5):461‑465. https://doi.org/10.1001/archpsyc.58.5.461
- Hemiplegia of early onset and the faculty of speech with special reference to the effects of hemispherectomy.Brain85(3):427‑460. https://doi.org/10.1093/brain/85.3.427
- A review of subtyping in autism and proposed dimensional classification model.Journal of Autism and Developmental Disorders31(4):411‑22. https://doi.org/10.1023/A:1010616719877
- Extensive piano practicing has regionally specific effects on white matter development.Nature Neuroscience8(9):1148‑1150. https://doi.org/10.1038/nn1516
- Critical periods during sensory development.Current Opinion in Neurobiology10(1):138‑145. https://doi.org/10.1016/S0959-4388(99)00047-1
- The critical period for alteration in cortical binocularity resulting from divergent and convergent strabismus.Developmental Brain Research2(2):181‑202. https://doi.org/10.1016/0165-3806(81)90031-6
- Effect of early institutionalization and foster care on long-term white matter development: a randomized clinical trial.JAMA Pediatrics169(3):211‑219. https://doi.org/10.1001/jamapediatrics.2014.3212
- Language recovery after left hemispherectomy in children with late-onset seizures.Annals of Neurology46(4):579‑586. https://doi.org/10.1002/1531-8249(199910)46:4<579::AID-ANA5>3.0.CO;2-K
- Memory, language and intellectual ability in low-functioning autism Memory in Autism.Memory in Autism. Cambridge: CUP.Cambridge. URL: http://eprints.bournemouth.ac.uk/9558/1/9780521862882c14_p268-290.pdf
- Sensitive periods for visual calibration of the auditory space map in the barn owl optic tectum.Journal of Neuroscience18(10):3929‑3942.
- Regional cerebral blood flow throughout the sleep-wake cycle.Brain120(7):1173‑1197. https://doi.org/10.1093/brain/120.7.1173
- Early development of abstract language knowledge: evidence from perception–production transfer of birth-language memory.Open Science4(1):160660‑160660.
- Functional Architecture and Spike Timing Properties of Corticofugal Projections From Rat Ventral Temporal Cortex.Journal of Neurophysiology100(1):327‑335. https://doi.org/10.1152/jn.90392.2008
- A review of BF Skinner's Verbal Behavior.Language35(1):26‑58. https://doi.org/10.2307/411334
- Ch1.Aspects of the Theory of Syntax.Massachusetts Institute of Technology,Cambridge, MA.
- Changes in cortical activity during mental rotation A mapping study using functional MRI.Brain119(1):89‑100. https://doi.org/10.1093/brain/119.1.89
- Genie: A Psycholinguistic Study of a Modern-Day "Wild Child".Academic Press.
- Dissociations between language and cognition: Cases and implications.Journal of Autism and Developmental Disorders11(1):15‑30. https://doi.org/10.1007/BF01531338
- Abnormal language acquisition and the modularity of language.Linguistics: the Cambridge survey2:96‑116.
- The case of Chelsea: The effects of late age at exposure to language on language performance and evidence for the modularity of language and mind.UCLA Working Papers in Linguistics18:115‑146.
- An update on the linguistic development of Genie.Georgetown University Round Table on Languages and Linguistics145‑157.
- Ictal Coprolalia: A Case Report and Review of Ictal Speech as a Localizing Feature in Epilepsy.Pediatric Neurology57:88‑90. https://doi.org/10.1016/j.pediatrneurol.2015.11.013
- The Descent of Man and Selection in Relation to Sex.John Murray,London.
- Arithmetic and the brain.Current Opinion in Neurobiology14(2):218‑224. https://doi.org/10.1016/j.conb.2004.03.008
- Right hemisphere specialization for mental rotation in normals and brain damaged subjects.Cortex26(2):177‑188. https://doi.org/10.1016/S0010-9452(13)80349-8
- Birdsong and human speech: common themes and mechanisms.Annual Review of Neuroscience22(1):567‑631. https://doi.org/10.1146/annurev.neuro.22.1.567
- Visual imagery and visual-spatial language: Enhanced imagery abilities in deaf and hearing ASL signers.Cognition46(2):139‑181. https://doi.org/10.1016/0010-0277(93)90017-P
- Interhemispheric synchronization of oscillatory neuronal responses in cat visual cortex.Science252(5009):1177. https://doi.org/10.1126/science.252.5009.1177
- Functional postnatal development of the rat primary visual cortex and the role of visual experience: dark rearing and monocular deprivation.Vision Research34(6):709‑720. https://doi.org/10.1016/0042-6989(94)90210-0
- The evolution of the language faculty: clarifications and implications.Cognition97(2):179‑210. https://doi.org/10.1016/j.cognition.2005.02.005
- Epidemiological surveys of autism and other pervasive developmental disorders: an update.Journal of Autism and Developmental Disorders33(4):365‑382. https://doi.org/10.1023/A:1025054610557
- Conduction Velocity Variations Minimiz Conduction Time DIfferences Among Rednal Ganglion Cell Axons.Science238.
- The development of language in genie: a case of language acquisition beyond the “critical period”.Brain and Language1(1):81‑107. https://doi.org/10.1016/0093-934X(74)90027-3
- The Prefrontal Cortex.Fourth Edition (4 edition).Academic Press
- Conceptual size representation in ventral visual cortex.Neuropsychologia81:198‑206. https://doi.org/10.1016/j.neuropsychologia.2015.12.029
- Lesion mapping of cognitive abilities linked to intelligence.Neuron61(5):681‑691. https://doi.org/10.1016/j.neuron.2009.01.026
- Separate visual pathways for perception and action.Trends in Neurosciences15(1):20‑25. https://doi.org/10.1016/0166-2236(92)90344-8
- Oscillatory responses in cat visual cortex exhibit inter-columnar synchronization which reflects global stimulus properties.Nature338(6213):334‑337. https://doi.org/10.1038/338334a0
- First-language acquisition in adolescence: Evidence for a critical period for verbal language development.Brain and Language63(2):237‑255. https://doi.org/10.1006/brln.1997.1943
- Neuroelectromagnetic correlates of perceptual closure processes.Journal of Neuroscience30(24):8342‑8352. https://doi.org/10.1523/JNEUROSCI.5434-09.2010
- Myelinization of the optic nerve and its dependence on visual function—a quantitative investigation in mice.Development11(1):255‑266.
- Selective right parietal lobe activation during mental rotation.Brain123(1):65‑73. https://doi.org/10.1093/brain/123.1.65
- Multiple sensitive periods in the development of the primate visual system.Science232:235‑239. https://doi.org/10.1126/science.3952507
- The faculty of language: What is it, who has it, and how did it evolve?science298(5598):1569‑1579. https://doi.org/10.1126/science.298.5598.1569
- The organization of behavior: A neuropsychological approach.John Wiley & Sons
- The relation between cognitive and motor dysfunction and motor imagery ability in patients with multiple sclerosis.Multiple Sclerosis Journal18(9):1303‑1309. https://doi.org/10.1177/1352458512437812
- Oscillatory synchronization in large-scale cortical networks predicts perception.Neuron69(2):387‑396. https://doi.org/10.1016/j.neuron.2010.12.027
- Dynamically modulated spike correlation in monkey inferior temporal cortex depending on the feature configuration within a whole object.Journal of Neuroscience25(44):10299‑10307. https://doi.org/10.1523/JNEUROSCI.3036-05.2005
- BDNF regulates the maturation of inhibition and the critical period of plasticity in mouse visual cortex.Cell98(6):739‑755. https://doi.org/10.1016/S0092-8674(00)81509-3
- To see but not to see: A case study of visual agnosia.Psychology Press
- Spatial and numerical abilities without a complete natural language.Neuropsychologia49(5):924‑936. https://doi.org/10.1016/j.neuropsychologia.2010.12.017
- The critical period for ocular dominance plasticity in the ferret’s visual cortex.Journal of Neuroscience19(16):6965‑6978.
- The wild boy of Aveyron (G. Humphrey & M. Humphrey, trans.).New York: Appleton-Century-Crofts. (Original work published 1806).
- Creation through contact; sign language emergence and sign language change in nicaragua.na
- Distinct cortical areas associated with native and second languages.Nature388(6638):171‑174. https://doi.org/10.1038/40623
- Myelination and isochronicity in neural networks.Frontiers in Neuroanatomy3https://doi.org/10.3389/neuro.05.012.2009
- Plasticity of auditory maps in the brain.Trends in Neurosciences14(1):31‑37. https://doi.org/10.1016/0166-2236(91)90181-S
- Monaural occlusion alters sound localization during a sensitive period in the barn owl.Journal of Neuroscience4(4):1001‑1011.
- The emergence of temporal language in Nicaraguan Sign Language.Cognition156:147‑163. https://doi.org/10.1016/j.cognition.2016.08.005
- A computational analysis of mental image generation: Evidence from functional dissociations in split-brain patients.Journal of Experimental Psychology: General114(3):311‑341. https://doi.org/10.1037/0096-3445.114.3.311
- Lateralization and the critical period.The Journal of the Acoustical Society of America52(1A):174‑174. https://doi.org/10.1121/1.1982096
- Role of myelination in the development of a uniform olivocerebellar conduction time.Journal of Neurophysiology89(4):2259‑2270. https://doi.org/10.1152/jn.00922.2002
- Neural correlates of superior intelligence: stronger recruitment of posterior parietal cortex.Neuroimage29(2):578‑586. https://doi.org/10.1016/j.neuroimage.2005.07.036
- The biological foundations of language.Hospital Practice2(12):59‑67. https://doi.org/10.1080/21548331.1967.11707799
- Stimulus overselectivity in autism: a review of research.Psychological Bulletin86(6):1236‑1254. https://doi.org/10.1037/0033-2909.86.6.1236
- Selective responding by autistic children to multiple sensory input.Journal of Abnormal Psychology77(3):211‑222. https://doi.org/10.1037/h0031015
- Does age of language acquisition affect the relation between American sign language and mental rotation?University of Minnesota
- Age of acquisition effects on mental rotation: evidence from Nicaraguan sign language.Proceedings of the 37th Boston University Conference on Language Development.
- The long-lasting advantage of learning sign language in childhood: Another look at the critical period for language acquisition.Journal of Memory and language30(4):486‑512. https://doi.org/10.1016/0749-596X(91)90018-F
- Echolalia's types in children with Autism Spectrum Disorder.Revista CEFAC17(6):2072‑2080.
- Chasing the mythical ten percent: parental hearing status of deaf and hard of hearing students in the United States.Sign Language Studies4(2):138‑163. https://doi.org/10.1353/sls.2004.0005
- Grammatical development in adolescent first-language learners.Linguistics41(4; ISSU 386):681‑722.
- Treatment of echolalia in individuals with autism spectrum disorder: a Systematic Review.Review Journal of Autism and Developmental Disorders3(1):82‑91. https://doi.org/10.1007/s40489-015-0067-4
- Maturational constraints on language learning.Cognitive Science14(1):11‑28. https://doi.org/10.1207/s15516709cog1401_2
- Representation of number in the brain.Annual Review of Neuroscience32:185‑208. https://doi.org/10.1146/annurev.neuro.051508.135550
- Profile of the sensitive period for monocular deprivation in kittens.Experimental Brain Research39(1):17‑21.
- Increased auditory cortical representation in musicians.Nature392(6678):811‑814. https://doi.org/10.1038/33918
- Uniform range of conduction times from the lateral amygdala to distributed perirhinal sites.Journal of Neurophysiology87(3):1213‑1221. https://doi.org/10.1152/jn.00623.2001
- Words and rules: The ingredients of language.Basic Books
- Stimulus overselectivity four decades later: A review of the literature and its implications for current research in autism spectrum disorder.Journal of Autism and Developmental Disorders40(11):1332‑1349. https://doi.org/10.1007/s10803-010-0990-2
- Children’s spatial thinking: does talk about the spatial world matter?Developmental science14(6):1417‑1430. https://doi.org/10.1111/j.1467-7687.2011.01088.x
- The cognitive outcome of hemispherectomy in 71 children.Epilepsia45(3):243‑254. https://doi.org/10.1111/j.0013-9580.2004.15303.x
- Evidence from an emerging sign language reveals that language supports spatial cognition.Proceedings of the National Academy of Sciences107(27):12116‑12120. https://doi.org/10.1073/pnas.0914044107
- Explicit encoding of multimodal percepts by single neurons in the human brain.Current Biology19(15):1308‑1313. https://doi.org/10.1016/j.cub.2009.06.060
- Sparse but not “grandmother-cell”coding in the medial temporal lobe.Trends in Cognitive Sciences12(3):87‑91. https://doi.org/10.1016/j.tics.2007.12.003
- The initial stages of first-language acquisition begun in adolescence: when late looks early.Journal of Child Language40(02):391‑414. https://doi.org/10.1017/S0305000911000535
- Progressive Matrices.Progressive Matrices.Oxford Psychologists Press
- Perception's shadow: long-distance synchronization of human brain activity.Nature397(6718):430‑430. https://doi.org/10.1038/17120
- Genie: A scientific tragedy.Harper Perennial.
- Change of conduction velocity by regional myelination yields constant latency irrespective of distance between thalamus and cortex.Proceedings of the National Academy of Sciences100(10):6174‑6179. https://doi.org/10.1073/pnas.0937380100
- Mental rotation and object categorization share a common network of prefrontal and dorsal and ventral regions of posterior cortex.Neuroimage35(3):1264‑1277. https://doi.org/10.1016/j.neuroimage.2007.01.012
- Diagnostic features of autism.Journal of Child Neurology3(1):57‑64. https://doi.org/10.1177/0883073888003001S11
- A human intracranial study of long-range oscillatory coherence across a frontal–occipital–hippocampal brain network during visual object processing.Proceedings of the National Academy of Sciences105(11):4399‑4404. https://doi.org/10.1073/pnas.0708418105
- The development of Nicaraguan Sign Language via the language acquisition process.Proceedings of the 19th Annual Boston University Conference on Language Development.
- Children creating language: How Nicaraguan Sign Language acquired a spatial grammar.Psychological Science12(4):323‑328. https://doi.org/10.1111/1467-9280.00359
- Children creating core properties of language: Evidence from an emerging sign language in Nicaragua.Science305(5691):1779‑1782. https://doi.org/10.1126/science.1100199
- Binding by synchrony.Scholarpedia2(12):1657. https://doi.org/10.4249/scholarpedia.1657
- Visual feature integration and the temporal correlation hypothesis.Annual Review of Neuroscience18(1):555‑586. https://doi.org/10.1146/annurev.ne.18.030195.003011
- The Neuropsychology of Dreams: A Clinico-anatomical atudy.Lawrence Erlbaum Associates
- Response of Schwann cells to action potentials in development.Science287(5461):2267‑2271. https://doi.org/10.1126/science.287.5461.2267
- Control of myelination by specific patterns of neural impulses.Journal of Neuroscience18(22):9303‑9311.
- Principles of Brain Evolution.Sinauer Associates
- Uniform olivocerebellar conduction time underlies Purkinje cell complex spike synchronicity in the rat cerebellum.The Journal of Physiology470:243. https://doi.org/10.1113/jphysiol.1993.sp019857
- Development of the equine brain motor system.Neurobiology (Budapest, Hungary)9(2):107‑135. https://doi.org/10.1556/Neurob.9.2001.2.4
- Response of the three main types of glial cells of cortex nad corpus callosum in rats handled during suckling or exposed to enriched, control and impoverished environments following weaning.Journal of Comparative Neurology172(2):247‑263. https://doi.org/10.1002/cne.901720205
- Myelination in rabbit optic nerves is accelerated by artificial eye opening.Neuroscience Letters16(3):235‑238. https://doi.org/10.1016/0304-3940(80)90003-8
- Childhood neglect is associated with reduced corpus callosum area.Biological Psychiatry56(2):80‑85. https://doi.org/10.1016/j.biopsych.2004.03.016
- A comparative study of myelination in precocial and altricial murid rodents.Developmental Brain Research43(1):139‑147. https://doi.org/10.1016/0165-3806(88)90159-9
- Mapping brain maturation.Focus.
- Neural synchrony in brain disorders: relevance for cognitive dysfunctions and pathophysiology.Neuron52(1):155‑168. https://doi.org/10.1016/j.neuron.2006.09.020
- Visual cognition and representation in birds and primates.Comparative vertebrate cognition.Springer
- The network substrate of confabulatory tendencies in Alzheimer's disease.Cortex87:69‑79. https://doi.org/10.1016/j.cortex.2016.08.015
- On the Origin of the Human Mind.2nd ed.MobileReference,Boston, Massachusetts.
- Mental synthesis involves the synchronization of independent neuronal ensembles.Research Ideas and Outcomes1:7642. https://doi.org/10.3897/rio.1.e7642
- Neurobiological mechanisms for nonverbal IQ tests: implications for instruction of nonverbal children with autism.Research Ideas and Outcomes3:e13239‑e13239. https://doi.org/10.3897/rio.3.e13239
- A system for relational reasoning in human prefrontal cortex.Psychological Science10(2):119‑125. https://doi.org/10.1111/1467-9280.00118
- Sparse Representation in the Human Medial Temporal Lobe.Journal of Neuroscience26(40):10232‑10234. https://doi.org/10.1523/jneurosci.2101-06.2006
- Wechsler intelligence scale for children.Psychological CorporationURL: http://psycnet.apa.org/psycinfo/1950-02930-000
- Speech and brain mechanisms.Princeton: Princeton University Press
- Prevalence of autism in a US metropolitan area.Jama289(1):49‑55. https://doi.org/10.1001/jama.289.1.49
- Neuroimaging studies of mental rotation: a meta-analysis and review.Journal of Cognitive Neuroscience20(1):1‑19. https://doi.org/10.1162/jocn.2008.20013
- Neural correlates of simple and complex mental calculation.Neuroimage13(2):314‑327. https://doi.org/10.1006/nimg.2000.0697
- By heart an fmri study of brain activation by poetry and prose.Journal of Consciousness Studies20(9-10):132‑158.
Supplementary material
This Power Point slide contains the content of the Figure 3.