|
Research Ideas and Outcomes : Workshop Report
|
|
Corresponding author: Adrian G Glover (a.glover@nhm.ac.uk)
Received: 16 Sep 2016 | Published: 16 Sep 2016
© 2016 Adrian G Glover, Thomas G Dahlgren, Sergio Taboada, Gordon Paterson, Helena Wiklund, Andrea Waeschenbach, Amber Cobley, Pedro Martínez, Stefanie Kaiser, Sarah Schnurr, Sahar Khodami, Uwe Raschka, Daniel Kersken, Heiko Stuckas, Lenaick Menot, Paulo Bonifacio, Ann Vanreusel, Lara Macheriotou, Marina Cunha, Ana Hilário, Clara Rodrigues, Ana Colaço, Pedro Ribeiro, Magdalena Błażewicz, Andrew J Gooday, Daniel OB Jones, David SM Billett, Aurélie Goineau, Diva J Amon, Craig R Smith, Tasnim Patel, Kirsty McQuaid, Ralph Spickermann, Stefan Brager.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Glover A, Dahlgren T, Taboada S, Paterson G, Wiklund H, Waeschenbach A, Cobley A, Martínez P, Kaiser S, Schnurr S, Khodami S, Raschka U, Kersken D, Stuckas H, Menot L, Bonifacio P, Vanreusel A, Macheriotou L, Cunha M, Hilário A, Rodrigues C, Colaço A, Ribeiro P, Błażewicz M, Gooday A, Jones D, Billett D, Goineau A, Amon D, Smith C, Patel T, McQuaid K, Spickermann R, Brager S (2016) The London Workshop on the Biogeography and Connectivity of the Clarion-Clipperton Zone. Research Ideas and Outcomes 2: e10528. doi: 10.3897/rio.2.e10528
|

|
Abstract
Background
Recent years have seen a rapid increase in survey and sampling expeditions to the Clarion-Clipperton Zone (CCZ) abyssal plain, a vast area of the central Pacific that is currently being actively explored for deep-sea minerals (ISA, 2016). Critical to the development of evidence-based environmental policy in the CCZ are data on the biogeography and connectivity of species at a CCZ-regional level.
New information
The London Workshop on the Biogeography and Connectivity of the CCZ was convened to support the integration and synthesis of data from European Union (EU) CCZ projects, supported by the EU Managing Impacts of Deep-Sea Resource Exploitation (MIDAS) and EU Joint Programming Initiative Healthy and Productive Seas and Oceans (JPI Oceans) projects. The London Workshop had three clear goals: (1) To explore, review and synthesise the latest molecular biogeography and connectivity data from across recent CCZ cruises from both contractor and academia-funded projects; (2) To develop complementary and collaborative institutional and program-based academic publication plans to avoid duplication of effort and ensure maximum collaborative impact; (3) To plan a joint synthetic data publication highlighting key results from a range of planned molecular biogeography/connectivity publications. 32 participants attended the workshop at the Natural History Museum in London from 10-12 May 2016. Presentations and discussions are summarised in this report covering (1) overviews of current CCZ environmental projects, (2) policy and industry perspectives, (3) synthesis of DNA taxonomy and biogeography studies, (4) summaries of the latest population genetic studies, (5) summaries of the latest broader morphological context, (6) an overview of publication and proposal plans to maximise collaborative opportunities and finally a series of workshop recommendations.
Keywords
Deep-sea biology, deep-sea mining, biodiversity, phylogeography, population genetics, European Union Framework 7, discussion meeting
Date and place
10-12 May 2016, Natural History Museum, London
List of participants
Same as author list.
Introduction
Recent years have seen a rapid increase in survey and sampling expeditions to the Clarion-Clipperton Zone (CCZ) abyssal plain, a vast area of the central Pacific that is currently being actively explored for deep-sea minerals (
Critical to the development of evidence-based environmental policy in the CCZ are data on the biogeography and connectivity of species at a CCZ-regional level. With this in mind, the London Workshop on the Biogeography and Connectivity of the CCZ was convened to support the integration and synthesis of data from European Union (EU) CCZ projects, supported by the EU Managing Impacts of Deep-Sea Resource Exploitation (MIDAS) and EU Joint Programming Initiative Healthy and Productive Seas and Oceans (JPI Oceans) projects, individual EU-based contractors and the Natural History Museum in London, a leading centre for marine biodiversity research.
The challenge of biogeography and connectivity in the CCZ
The Clarion-Clipperton Zone is so called as it lies between the Clarion and Clipperton Fracture Zones, topographical highs that extend longitudinally across almost the entire Pacific (Fig.

Exploration contract areas for polymetallic nodules in the Clarion-Clipperton Zone, central Pacific Ocean. Areas of Particular Environmental Interest (APEIs) numbered according to the latest International Seabed Authority data (Source: Stefan Brager, ISA), see section in this report: ‘Key Outcomes and Discussion’. Image credit: International Seabed Authority, 2015.
Conducting regional-level studies of the biogeography and connectivity of the CCZ is an immense challenge for several reasons: e.g., (1) the great depth and long distances to home ports, (2) the great physical heterogeneity of the region (the CCZ is not a homogenous abyssal plain of mud, it is a region of 6 million km2 with a bathymetric variation of at least 1200 m and punctuated by many thousands of seamounts) and (3) there are almost no data on habitat characteristics such as food availability or prevailing oceanographic currents. However, these are only minor issues compared with the greatest problem: the almost complete lack of taxonomic synthesis or standardisation across the region based either on traditional morphological data or modern DNA (
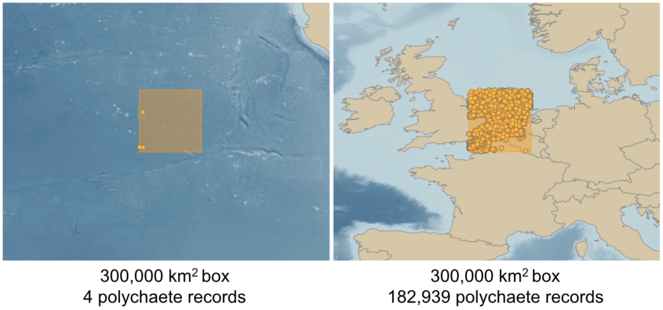
There is an almost complete absence of published, databased benthic species records from the Clarion-Clipperton Zone (CCZ), despite over 30 years of intensive oceanographic and geological research in the area. Here illustrated as an example are the current species records for a 300,000 km2 (5°) box centred on the North Sea and Eastern Channel in Europe, where the Ocean Biogeographic Information System (
Despite these problems, new data are now emerging from both academia and contractor-led CCZ programs that utilise genetic data in the form of short DNA sequences from typical invertebrate markers such as cytochrome oxidase I mitochondrial gene (COI), 16S mitochondrial ribosomal RNA coding genes and 18S or 28S nuclear genes. Recent publications from the BGR-led and UKSRL-led projects have made these data available on the open data repository NCBI GenBank (

Recent publications on the Clarion-Clipperton Zone are making imagery and genetic data available in public databases that will allow future workers access to these data and the voucher materials, for regional-level syntheses and further DNA sequencing if needed. Examples are recent taxonomic data papers on the Echinodermata (
The London Workshop was expressly designed to try and overcome these challenges by bringing together researchers from a range of EU projects working on molecular data from the CCZ to share findings and plan future publications and synthetic activities.
Aims of the workshop
Goals
The London Workshop had three clear goals:
- To explore, review and synthesise the latest molecular biogeography and connectivity data from across recent CCZ cruises from both contractor and academia-funded projects
- To develop complementary and collaborative institutional and program-based academic publication plans to avoid duplication of effort and ensure maximum collaborative impact
- To plan a joint synthetic data publication highlighting key results from a range of planned molecular biogeography/connectivity publications
During discussions regarding the workshop it also became apparent that it was important to review historical knowledge of CCZ biogeography based on morphological data, new data from visual survey tools and taxonomic descriptions alongside the new molecular data – this is discussed in more detail in section “Workshop Recommendations” below.
London Workshop: The Agenda
The agenda is presented here as planned in order to provide transparency of discussion and a useful guide to those planning future similar workshops. Summaries of talks and discussion are provided in the following section “Key outcomes and discussions”.
Tuesday 10 May 2016
1200 Lunch in the Natural History Museum North Hall
1300 Adrian Glover: Welcome, meeting logistics and participant introductions
1330 Adrian Glover: Overview of meeting goals and agenda, discussion
1400 Session 1: Project Overviews
The purpose of Session 1 was to introduce each relevant EU CCZ project from the point of view of molecular biogeography and connectivity data.
1400 Pedro Martinez: Overview of the German exploration claim (BGR) recent cruises and data collected
1420 Thomas Dahlgren & Pedro Martinez: Overview of the JPI-Oceans cruise program and data collected
1440 Ann Vanreusel: Overview of biological and environmental sampling in the GSR (Belgian) exploration area
1500 Lenaick Menot: Overview of the IFREMER (France) environmental studies
1520 Tea/coffee break
1540 Daniel Jones & Sergi Taboada: Overview of the NERC-MIDAS RRS James Cook JC120 cruise to APEI-6, including overview of molecular collecting
1600 Adrian Glover: Overview of the ABYSSLINE (UK Seabed Resources Ltd (UKSRL)) data on molecular biogeography and connectivity
1620 Session 2: Policy and Industry Perspectives
1620 Stefan Brager: The policy perspective: The International Seabed Authority and environmental management of the CCZ
1640 Ralph Spickermann: The industry perspective: Manganese nodules in the Pacific ocean, path from exploration to exploitation
1730 End of Day 1: Icebreaker social at the Hereford Arms pub, South Kensington.
Wednesday 11 May 2016
0900 Arrival and summary of Day 1, overview of Day 2 agenda.
0910 Thomas Dahlgren: Discussion Starter: Recent molecular connectivity papers in the deep sea, lessons learned
0930 Session 3: DNA taxonomy and biogeography within CCZ exploration claim areas
The purpose of Session 3 was to review new data on DNA taxonomy from the CCZ, that is the formal or informal description of species using DNA barcodes, and the examination of the distribution of those species within exploration areas.
0930 Pedro Martinez: Molecular taxonomy and biogeography within the German (BGR) exploration area
0950 Helena Wiklund: Molecular taxonomy and biogeography within the UKSRL, Ocean Mineral Singapore (OMS) and Area of Particular Environmental Interest (APEI) #6.
1010 Paulo Bonifacio: Diversity and distribution patterns of Polynoidae (Annelida) across the CCZ
1030 Tea/coffee break
1050 Lara Macheriotou: Deep-sea Nematoda of the CCZ – preliminary insights
1110 Andrew Gooday: Foraminifera species diversity within the the UKSRL and OMS exploration areas
1130 Magdalena Błażewicz: Abyssal Tanaidacea – (not) stunning cryptic diversity
1150 Andrea Waeschenbach: Abyssal Bryozoa – first results from UKSRL area and comments on CCZ projects
1210 Discussion session
1300 Lunch break
1400 Session 4: Intra-specific population connectivity in target taxa across the CCZ
The purpose of Session 4 was to review new data on population genetics and connectivity within populations, for which new data are starting to emerge from recent CCZ projects. These data are extremely limited but have the potential to offer statistically-robust estimates of connectivity for the first time. Typically, ‘target’ taxa are chosen that have large enough sample sizes.
1400 Heiko Stuckas: Population structure and demography in deep-sea species using molecular markers
1420 Thomas Dahlgren: Population structure in target taxa (Annelida, Mollusca and Echinodermata) from the UKSRL, OMS and APEI-6 areas
1440 Pedro Martinez: Population structure in Ophiuroidea from the CCZ
1500 Sarah Schnurr: Phylogeographic patterns in Isopoda from the JPI Oceans project
1520 Uwe Raschka: Phylogeographic patterns in Harpacticoida from the JPI Oceans project
1540 Tea/coffee break
1600 Sergi Taboada: Molecular connectivity of the sponge Tethyida sp. nov. within the CCZ using microsatellites
1620 Pedro Ribeiro: Using RAD sequencing (RADseq) data to investigate population genetic structure in the deep sea
1640 Session 5: The broader biogeographic context based on morphology and molecules
The purpose of the final data session was to review data on the broader biogeographic context of the CCZ, particularly based on historical morphological data coupled to new molecular studies. In particular, what new identified species records of actual described species can tell us about ranges at what scales.
1640 Adrian Glover: Annelida, Mollusca, Echinodermata: Actual species identifications from the CCZ using historical morphological data: some examples
1700 Daniel Kersken: Porifera of the CCZ
1720 Andrew Gooday: Foraminifera, the broader biogeographic context
1740 Gordon Paterson: The problem and challenge of rarity in the abyss
1800 Discussion
1900 Workshop Dinner, Ognisko Restaurant, South Kensington.
Thursday 12 May 2016
0800 Principal Investigator break-out session to agree on conceptual framework for a synthesis paper
0845 Workshop Photograph
0900 Summary of Day 2, discussion and overview of Day 3 agenda
0930 Session 6: Summary tables and data synthesis session
The purpose of the final summary session was to review what data are currently available (published or made available on open databases), what data are in the process of being published, and what data could be shared in collaborative, synthetic publications or reports.
0930 Session 6 discussion and production of summary table of planned publications
1030 Tea/coffee break
1100 Continuation of summary table production
1200 Round-table discussions on future cruises and grant proposals for report
1230 General workshop recommendations – summary table for report
1300 Workshop close
Key outcomes and discussions
Participant composition and overview of workshop goals
The workshop opened with participant introductions and a discussion of the proposed agenda. 32 participants attended the workshop, based on invitation sent to the MIDAS, JPI Oceans and EU-led contractor programs. The meeting was also advertised via the MIDAS news feed and the INDEEP email alert. The workshop was over-subscribed, with an initial capacity of 25, but the venue was expanded to accommodate the 32 accepted invitees. The meeting was maintained within the EU-led projects in order to achieve a manageable size to facilitate discussion, although it was noted that the potential number of attendees could have been much higher if the workshop had been expanded to include non-EU contractors and Sponsoring State research programs. The workshop participants agreed that this could be the subject of a future workshop or specialist session at an international meeting (see Workshop Recommendations).
23 of the 32 attendees had specialist taxonomic knowledge in particular phyla (Fig.
In terms of professional status, the meeting was dominated by Principal Investigators (56%), then post-graduate students (25%) and Post-docs (19%) (Fig.

Composition of participants in the London Workshop on the Biogeography and Connectivity of the Clarion-Clipperton Zone, based on taxon expertise (only estimated in 23 participants), professional status, representing nation and gender (from 32 participants).
Following participant introductions, the workshop commenced with an overview from Adrian Glover on the workshop goals. Glover pointed out the rapid growth in exploration activity in the CCZ (e.g. the number of exploration contracts has more than doubled in the last 10 years (
There was agreement on the overarching goals of the workshop:
- To explore, review and synthesise the latest molecular biogeography and connectivity data from across recent CCZ cruises from both contractor and academia-funded projects
- To develop complementary and collaborative institutional and program-based academic publication plans to avoid duplication of effort and ensure maximum collaborative impact
- To plan a joint synthetic data publication highlighting key results from a range of planned molecular biogeography/connectivity publications
There was also general agreement on the need for a synthesis paper, although the delivery mechanism for this third goal remained unclear at the start of the workshop, and is discussed further below.
Session 1: An overview of recent CCZ cruises and projects of relevance to molecular biogeography and connectivity
Extensive collection for DNA taxonomy and biogeographic analysis first started in the CCZ in 2003-2004 as part of the Kaplan project (
At the London Workshop, Pedro Martinez provided an overview of the recent German (BGR) environmental work in their exploration claim area. Martinez explained that BGR have two claim areas, one in the eastern CCZ and one in the western CCZ, and the site in the eastern CCZ has been much more intensively explored (Fig.

The BGR-Germany polymetallic nodule exploration claim area in the eastern CCZ, illustrating the two areas in which survey work has concentrated in recent years - the proposed preservation reference area (PRA) and impact reference area (IRA). Image source: Federal Institute for Geosciences and Natural Resources (BGR, Germany) and Pedro Martinez, Senckenberg Institute (
Pedro Martinez also outlined the JPI Oceans program aboard the R/V Sonne (

Outline of the JPI Oceans ‘SO-239’ research cruise aboard the R/V Sonne 10 March – 30 April 2015 to the eastern CCZ (Image from Short Cruise Report: Martinez, 2015).
Discussion at the Workshop about the JPI Oceans project was mainly focussed on the data from the tracks and the impacts on connectivity. For example, David Billett raised the question of the distance at which visible (sedimentation on nodules) impacts of the old OMCO tracks were still visible. Daniel Jones pointed out that a significant portion of their work with Autonomous Underwater Vehicles (AUVs) is focussed on developing high-resolution mapping of the impacts from disturbances. Discussion moved on to the importance of understanding the hydrodynamics of these sites and the potential role of larval dispersal and larval biology, raised by Ana Hilario. Marina Cunha pointed out that a more precise working definition of ‘connectivity’ would be useful, with recent reviews published in the literature (see Workshop Recommendations).
Environmental sampling from the Belgian (GSR) contracted area was reviewed by Ann Vanreusel. Sampling has taken place in the Belgian area in the eastern CCZ (between the German and French areas) during GSR cruises in 2014, 2015 and as part of SO-239 in 2015. Molecular samples were obtained from 'multiple core' and 'box core' samples for meiofauna (DESS preserved) and macrofauna (ethanol preserved) in samples from three areas within the Belgian claim (Fig.

The GSR-Belgium polymetallic-nodule exploration claim areas in the eastern CCZ, illustrating the three areas in which survey work has concentrated, and the sampling stations in which molecular data were obtained from GSR cruise in 2015 and the JPI Oceans SO-239 cruise in 2015. Image source: Ann Vanreusel, University of Ghent.
Lénaick Menot presented an overview of cruises and studies carried out by Ifremer (France) in the Clarion-Clipperton Zone. Ifremer was the lead on the ‘Nodinaut’ cruise onboard RV L’Atalante with the submersible Nautile in 2004 and the ‘BIONOD’ cruise onboard RV L’Atalante in 2012, in which both cruises contributed significant samples for molecular biology. The Nodinaut cruise was itself a major contribution to the Kaplan project. Two French mining exploration areas have been sampled in the center and to the west of the CCZ. The BIONOD cruise resulted from collaboration between Ifremer and BGR in which both a German claim in the eastern CCZ and a French claim in the central CCZ were visited. During both cruises, meiobenthic and macrobenthic communities were sampled. Kaplan samples during the Nodinaut cruise and epibenthic sledge samples during the BIONOD cruise were preserved in cold ethanol to facilitate DNA barcoding of the species. Results of the BIONOD cruise allowed publication of the first biodiversity assessment of polychaetes and isopods based on a barcoding approach in the CCZ in 2015 (
Daniel Jones presented an overview of the recent MIDAS cruise aboard RRS James Cook (JC120), co-funded by the Natural Environment Research Council (NERC) National Capability funding. The cruise program was dedicated to the characterisation of the APEI to the north-east of the CCZ region, formerly known as ‘APEI-4’. It is important to note that this has recently been re-labelled as APEI-6, and the new APEI numbering system is illustrated in Fig.
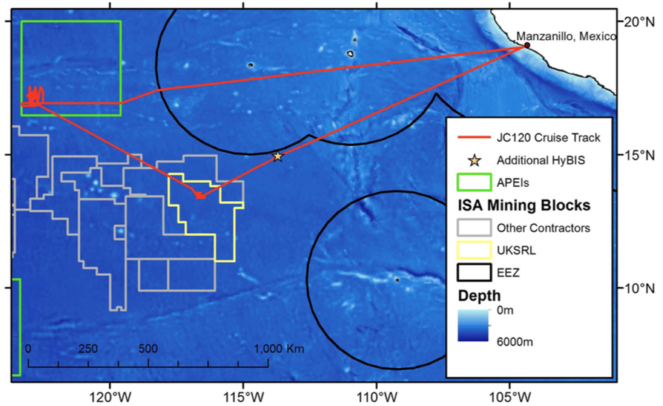
The eastern CCZ, illustrating the cruise track of the RRS James Cook MIDAS JC-120 cruise to sample APEI-6 (green box) and a short sampling station in the northern sector of UK-1 (yellow box). Image source: Daniel Jones, National Oceanography Centre, UK.
The macrofaunal and megafaunal DNA component of the JC120 samples is being funded directly by the MIDAS through Work Package 4 (Task 4.1) and being led by Sergi Taboada, Gordon Paterson and Adrian Glover in collaboration with Daniel Jones. Taboada provided an overview of the relevant data that is being provided. Samples were obtained from box core, mega core, Hybis ROV, Agassiz trawl and amphipod traps. 462 samples were taken for DNA analysis, 90% of them from the APEI-6 and 10% from UK-1. 81 species were determined based on barcoding analysis, 15 of them annelids (Fig.

The DNA taxonomy field pipeline for the Managing Impacts of Deep Sea Resource Exploitation (MIDAS) cruise aboard RRS James Cook (JC120). Image source: Sergi Taboada, Natural History Museum, UK.
Adrian Glover provided the final project overview of the day with a discussion on the molecular connectivity and biogeography parts of the ABYSSLINE project, funded by UKSRL. This is a contractor baseline survey that is being undertaken through a collaborative partnership with a globally-distributed academic partner network, coordinated and led by Craig Smith at the University of Hawaii. Partners in the UK include the National Oceanography Centre and the Natural History Museum. The project has been running since mid-2013, with cruises taking place in October 2013 (R/V Melville) and February-March 2015 (R/V Thomas G Thompson). The sampling design consists of a series of 30x30km boxes within which randomised sampling takes place using a wide range of equipment, the most relevant to the DNA work being the epibenthic sledge, 'box core', 'multi core' and ROV (
Summary of Clarion-Clipperton Zone (CCZ) EU-contractor/academia led projects with a significant molecular biology (DNA taxonomy or population genetics) component
|
Project Name |
Time Period |
Cruises & Year |
Principal DNA Work |
Key DNA Publications |
|
KAPLAN |
2003-2004 |
R/V New Horizon 2003; R/V Umitaka-Maru 2004; R/V L'Atalante 2004 |
Macro- and meiofauna DNA taxonomy |
Smith et al 2008 |
|
BGR (Germany) |
2010- ongoing |
R/V Sonne and R/V Kilo Moana; ~Annual since 2010 |
Mega-, macro- and meiofauna DNA taxonomy & connectivity |
Janssen et al 2015 |
|
JPI-Oceans |
2015-ongoing |
R/V Sonne 2015 |
Mega-, macro- and meiofauna DNA taxonomy |
In progress |
|
GSR (Belgium) |
2014-ongoing |
2014, 2015 (JPI-Oceans) |
Meiofauna |
In progress |
|
Ifremer (France) |
2004-ongoing |
R/V L'Atalante 2004; R/V L'Atalante 2012 |
Mega-, macro- and meiofauna DNA taxonomy |
Janssen et al 2015 |
|
MIDAS (NOC-NERC) |
2015 |
RRS James Cook JC120 2015 |
Mega-, macrofauna DNA taxonomy & connectivity |
In progress |
|
ABYSSLINE (UKSRL) |
2013-ongoing |
R/V Melville 2013; R/V Thomas G Thompson 2015 |
Mega-, macro- meio- and microbial fauna DNA taxonomy & connectivity |
Glover et al 2015; Glover et al 2016; Dahlgren et al 2016 |

The UK Seabed Resources Ltd (UKSRL) Abyssal Baseline (ABYSSLINE) survey area in the UK-1 exploration area in the eastern CCZ, with the UK-1 and Ocean Mineral Singapore (OMS) reserved areas highlighted. The survey design consists of replicated 30x30km boxes within the exploration area within which randomised sampling using a range of equipment is undertaken (Smith et al 2013). Image source: Craig R Smith, ABYSSLINE Project.
Session 2: The policy and industry perspective
Although primarily a data-discussion workshop, it was thought important to include reasonable time on policy and industry perspectives in order to direct discussion and to provide direct two-way dialogue with the development of the regulatory framework at the ISA. With this in mind, Stefan Brager (ISA) and Ralph Spickermann (UKSRL) were invited to the workshop to contribute from these perspectives.
Stefan Brager presented the policy background for contractor investments into the study of biogeography and connectivity in the CCZ. To protect the Common Heritage of Mankind, the Law of the Sea has mandated the ISA to create rules and regulations to ensure protection of the marine environment during deep-sea mining. The ‘Environmental Management Plan for the Clarion-Clipperton Zone’ (ISBA/17/LTC/7, ISA 2011) is such a binding document. It was adopted in 2012 and is currently up for review. Besides creating the nine existing Areas of Particular Environmental Interest (APEI) (Fig.
Dr Brager’s presentation resulted in several discussion points being raised at the London Workshop. There was a short discussion on how aware (or not) the academic scientific community is with regard to the various regulatory developments, in particular important dates for reviews, milestones, adoptions of regulations and changes etc. Examples are the need for review of the APEIs, the deadline for that, and the need for draft exploitation guidelines, due in July 2016. A policy brief for the academic community may be of use (see Workshop Recommendations). Brager challenged the scientists in the room to answer if they felt the goals of the CCZ EMP (
Following this discussion, Ralph Spickermann (UKSRL) addressed the workshop from the industry perspective. His presentation outlined the path from exploration to exploitation from the contractor point of view. Spickermann pointed out that seabed minerals are essentially a capital-intensive new industry, with two key business enablers being the regulatory framework and the environmental responsibility. From an industry perspective, the future exploitation regulations (currently under discussion at the ISA) must be commercially viable and environmentally sustainable, stable and predictable for an infant industry, reflecting of the technical risk relative to terrestrial ventures and with a simple royalty structure. In terms of resource certification (i.e how much of a resource there actually is), this can follow already well-established protocols such as the Canadian NI 43-101 or equivalent. Spickermann finished with a brief overview of the ABYSSLINE consortium environmental baseline work, highlighting the cruises (mentioned by Adrian Glover earlier in his overview) and the broad range of studies being carried out, and emphasising that the value of sample richness is only realised upon analysis and publication, not just collection. This latter point is broadly supported by the science community and particularly relevant to the CCZ where there has been a long history of biological sample taking without appropriate funding models to work up the samples (see Workshop Recommendations).
Session 3: Synthesis of DNA taxonomy and biogeography within CCZ exploration claim areas
The third session of the London Workshop addressed what has been the main focus of attention in recent DNA work in the CCZ: what we term in this report ‘DNA taxonomy’. There is often a mixture of use of the terms DNA taxonomy and DNA barcoding in the literature. In its strictest sense, DNA barcoding refers to the identification of a species by sequencing a known marker gene, often the COI marker, and comparing this against known databases or libraries (Hebert et al 2003). In contrast, DNA taxonomy (at least as we define it) is the creation of that database or library. This is particular important in the deep sea, where we have seen in recent CCZ examples (
The session started with Pedro Martinez highlighting results from the joint BGR-Ifremer efforts to the German and French exploration areas. Data from the BGR cruise SO205 in 2010 and the BIONOD Ifremer cruise in 2012 have been published in

Analysis reproduced from
Helena Wiklund provided an overview of the DNA taxonomy work on the UKSRL ABYSSLINE project. The project team (Wiklund, Adrian Glover and Thomas Dahlgren) are working on a collection of 3312 individually databased and photographed megafaunal and macrofaunal specimens (excluding Crustacea) from the two ABYSSLINE cruises conducted in 2013 and 2015 (Fig.

Selected live images of specimens recovered from the UK Seabed Resources Ltd (UKSRL) Abyssal Baseline (ABYSSLINE) research cruise to the eastern Clarion-Clipperton Zone, February-March 2015. Image credit: Adrian Glover, Thomas Dahlgren, Helena Wiklund.
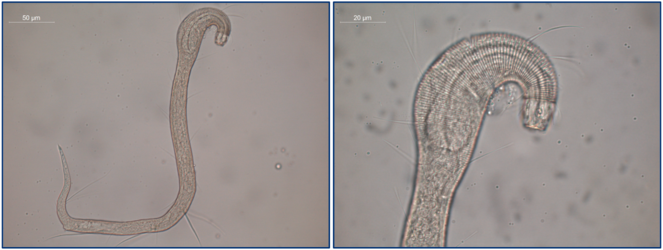
Nematoda of the Clarion-Clipperton Zone, recovered from abyssal sediments during the JPI Oceans ‘SO-239’ cruise aboard the R/V Sonne in February-March 2015. Image credit: Lara Macheriotou, University of Ghent.
At the London Workshop, Wiklund outlined some of the new data from the second ABYSSLINE cruise (AB02) and in addition analysed these data alongside new unpublished data from the MIDAS-JC120 cruise (see above). In general, some evidence for broad species ranges was found in a range of taxa when regional-level data was included from the UK-1, OMS, Ifremer and BGR exploration contract areas. However, it was pointed out that it is only possible to demonstrate presence, not absence, given the likelihood of under-sampling. This is a common problem thread that runs through much of the regional-level analyses being undertaken at present (see Workshop Recommendations). Discussion focussed on this, and in addition Lenaick Menot pointed out that it would be very interesting to focus some efforts on these broadly-distributed species to examine their functional traits, for example. Wiklund pointed out that as new data accumulate, new ‘target taxa’ emerge such as the pycnogonids, which seem to be quite broadly-distributed. Adrian Glover pointed out that we need to at some point stop thinking about the CCZ in terms of the contract boxes, and start to think more broadly about the region and the drivers of heterogeneity in general.
Paulo Bonifácio, together with Lenaick Menot and Lenka Neal, presented new data on the diversity, distribution and connectivity of polynoid worms across the CCFZ. The study aims to describe new species of deep-sea polynoids using morphology complemented with molecular data, evaluating monophyly of the subfamily Macellicephalinae and examining the genetic connectivity for widely distributed species found among different sampled areas (BGR, IOM, GSR, Ifremer and APEI#3). Samples were collected using an epibenthic sledge, box core (0.25m2) and ROV from the French and German cruise programs. Preliminary results suggested the presence of 40 morphotypes of Polynoidae, with the subfamily Macellicephalinae being the most abundant and species rich. Further, Bathyfauvelia sp. A, was found in 4 of the 5 studied areas indicating no evidence for a biogeographic barrier and the connectivity appears to be high between the areas at least 100 km apart from each other. Bonifácio concluded that their study exemplifies the need for a dual approach in taxonomy, phylogeny and connectivity studies, i.e combining morphological and genetic data.
Lara Macheriotou presented molecular barcoding work completed thus far relevant to the biodiversity and population connectivity of free-living Nematodes of the CCZ. These samples were collected during the SO-239 (JPI Ocean EcoResponse, March-April 2015) cruise which visited four contractor licences (BGR, IFREMER, IOM, GSR) and APEI#3, as well as the GSR-led campaign (GSRNOD15A, September-October 2015) to their respective claim. Despite being very preliminary results, these data are providing the first initial insights to the nematofauna of the CCZ based on DNA. Most prominent was the large discrepancy in generic diversity of Nematodes collected using methodologies pertaining to meiofaunal versus macrofaunal taxa, the latter being significantly lower than the former. Data specific to the most abundant and widespread genera (Halalaimus, Phanodermopsis) confirm these are cosmopolitan and tentatively point to the possibility of an endemic deep-sea species from genus Halalaimus. Discussion of the Macheriotou presentation focussed on a couple of issues: firstly the reduced success rate with COI relative to 18S (noted by many other researchers across a range of taxa) and secondly the interesting observations of the large numbers of macrofaunal-sized nematodes found, particularly in the box core samples.
Andrew Gooday presented an overview of the UKSRL ABYSSLINE studies on benthic foraminiferal diversity with a focus on the new data based on DNA. Samples were obtained from all the ABYSSLINE survey boxes (see above). Morphological analyses of only eight preserved coretop samples revealed very high levels of species richness, with over 500 morphospecies recognised and many others recorded from shipboard sorting (Fig.

Xenophyophores of the Clarion-Clipperton Zone, recovered from abyssal sediments during the two UKSRL Abyssal Baseline (ABYSSLINE) cruises aboard R/V Melville and R/V Thomas G Thompson in 2013 and 2015. Image credit: Andrew Gooday and Aurélie Goineau, National Oceanography Centre, UK.
Magda Błażewicz outlined the latest data on the tanaidaceans, a quite common crustacean found in abyssal sediments that lives in self-constructed tubes. Błażewicz is working on samples from the eastern CCZ (JPI Oceans project) together with material from another west Pacific project, the KuramBIO program. Generally, tanaidaceans are thought to have quite restricted distributions owing to their virtually sessile lifestyle (building the tubes) and reproductive strategy that lack planktonic stages throughout their lifetimes. Out of 67 specimens of the genus Pseudotanais (Fig. 15), apparently the most abundant tanaidacean genus from the CCZ, DNA (COI) was obtained from 41. She has emphasised that success ratio in extracting and amplyfing DNA was substantially higher from fresh material (extraction done onboard). Applying automatic procedures to delimit species (e.g. ABGD, GYMS) it has been demonstrated that Pseudotanais is represented by 12 species in the CCZ and that the genus itself is not monophyletic. The molecular results are confirmed by morphological analyses. It was observed that only two species were broadly-distributed in the CCZ (between IOM and BGR) while there was no taxon in common across both the CCZ and KuramBio. Data are in preparation for a publication.
Figure
Andrea Waeschenbach provided an overview of deep-sea and abyssal Bryozoa, including some preliminary data from some of the first DNA studies to be conducted in the CCZ as part of the UKSRL ABYSSLINE project. A general feature of deep-sea bryozoans is they are usually attached to hard substrates, with colonies raised above the substrate (Fig.

A new species of cyclostome bryozoan from the eastern CCZ UKSRL ABYSSLINE project. Image credit: Adrian Glover, Thomas Dahlgren and Helena Wiklund. Identification by Dennis Gordon via Andrea Waeschenbach.
Sarah Schnurr presented an overview of the DNA taxonomy and biogeography in the isopods from the CCZ, which are dominated by the Asellota. These are generally understood to be brooding species with limited powers of dispersal, with distribution and gene flow dependent on passive and active migration of adults. Schnurr presented data from the JPI Oceans SO-239 and SO-242-1 cruises that took place in 2015 to the eastern CCZ (SO-239, see above) and the east Peru Basin DISCOL area (SO-242-1, see above, Fig.

Sampling stations in the eastern CCZ and east Peru Basin for the joint JPI Oceans cruises SO-239 and SO-242 utilised for phylogeographic studies of the asellote isopods being conducted by Sarah Schnurr, Senckenberg Institute. Image source: Sarah Schnurr, Senckenberg Institute.
Tasnim Patel spoke about her work on Amphipoda (Table
Summary of Clarion-Clipperton Zone (CCZ) projects studying DNA taxonomy and biogeography with key parameters and publications.
| Taxon Group | Current lead scientist(s) | Cruise/area | Markers used | Key Publications |
| Isopoda, Polychaeta | P Martinez, A Janssen | BGR, Ifremer | COI | Janssen et al 2015 |
| Polychaeta, Mollusca, Cnidaria, Echinodermata & misc. | H Wiklund, A Glover, T Dahlgren, J Taylor, S Taboada | UK-1, OMS, APEI-6 | COI, 16S, 18S, 28S | Glover et al 2016; Dahlgren et al 2016 |
| Polynoidae | P Bonifácio, L Menot, L Neal | BGR, IOM, GSR, Ifremer, APEI-3 | COI, 16S | In preparation |
| Nematoda | L Macheriotou | BGR, Ifremer, GSR, IOM, APEI-3 | COI, 18S | In preparation |
| Foraminifera | A Gooday | UK-1, OMS | 18S | In preparation |
| Tanaidacea | M Blazewicz | eastern CCZ | COI | In preparation |
| Bryozoa | A Waeschenbach, P Taylor, A Glover, H Wiklund, T Dahlgren | UK-1, OMS, APEI-6 | COI, 16S, 18S, 28S | In preparation |
| Amphipoda | T Patel | BGR, Ifremer, GSR, IOM, APEI-3, DISCOL | COI, and RADseq | In preparation |
| Isopoda (JPI Oceans) | S Schnurr | BGR, Ifremer, GSR, IOM, APEI-3, DISCOL | COI, 16S, 18S | In preparation |
Summary of Clarion-Clipperton Zone (CCZ) projects studying population genetics with key parameters and publications.
| Target Taxa for Pop Gen | Current lead scientist(s) | Cruise/area | Markers used | Key Publications |
| Paralacydonia sp, Sigalionidae sp, Bathyglycinde sp, misc. Polychaeta, Macrostylis sp, Desmosomatidae sp, Eugerdella sp | H Stuckas, P Martinez, A Janssen | BGR | COI | In preparation |
| Bathyglycinde profunda, Paralacydonia sp, Lumbrinerides sp, Maldanidae, Nereididae, Ophiuroidea, Mollusca (Nucula sp) | T Dahlgren, A Glover, H Wiklund | UK-1, OMS, APEI-6 | COI | In preparation |
| Ophiuroidea | P Martinez | BGR, IOM, GSR, Ifremer, APEI-3, DISCOL | COI | In preparation |
| Pseudotachidius bipartitus | U Raschka | BGR, Ifremer, GSR, IOM, APEI-3 | COI | In preparation |
| Timea or Hemiasterella sp | S Taboada, A Glover, G Paterson, H Wiklund, T Dahlgren | UK-1, APEI-6 | microsatellites and NGS | In preparation |
Session 4: Synthesis of intra-specific population connectivity in target taxa across the CCZ
The second major data session of the London Workshop was focussed on individual target taxa, in which detailed population genetic analyses have been carried out. These data are extremely new and nothing has yet been published from the CCZ. In most instances, researchers are investigating a small number of taxa in which sufficient sample numbers are available from individual locations, and analysis can be undertaken using rapidly-evolving markers such as COI on the genetic heterogeneity of the populations, measured by analysis of haplotype diversity and analysis of demographic patterns. These analyses can also sometimes be used to infer patterns of speciation (
Thomas Dahlgren opened the session by providing a historical overview of population connectivity studies in the deep sea. In the early period of deep-sea exploration, the deep sea was considered a broadly stable and homogenous environment that would favour low genetic diversity (e.g.
The Dahlgren presentation at the London Workshop opened up discussion, chaired by Adrian Glover and some key unanswered questions relevant to the CCZ were proposed (see Workshop Recommendations):
- How do we go from indirect evidence of connectivity based on genetics to direct evidence based on functional traits and larval biology?
- What oceanographic data (currents, models) are available to support indirect evidence of connectivity?
- What genetic markers should we use to infer connectivity, and how does this vary between taxonomic groups?
Gordon Paterson raised the point that at what point do cryptic species matter if they perform the same ecological function? Ann Vanreusel pointed out that there are studies that show cryptic species are important at a functional level, and Adrian Glover pointed out that his lab are currently working on looking at functional differences measured by stable isotope analysis in cryptic polychaete species (PhD Student Madeleine Brasier). David Billett raised an interesting question as to whether we should be looking at also north-south patterns across the CCZ as well as east-west, and the effects of depth. Andrew Gooday noted that another variable that may structure populations is the calcium-carbonate compensation depth (CCD).
Heiko Stuckas provided an overview of haplotype diversity methods applied to deep-sea taxa (Fig.

An explanation of the study of haplotype diversity (Hd). Hd is measured by calculating the frequency of different haplotypes (genetically distinct sets of genes defined here by single nucleotide polymorphisms). The measurement of Hd and the frequency distribution of haplotypes can provide information on the genetic diversity of the population, the presence of speciation and demographic patterns. Image: Heiko Stuckas, Senckenberg Institute.
Thomas Dahlgren continued the population session with an overview of the study of ‘target taxa’ from the UKSRL ABYSSLINE project (Fig.

An example of a ‘target taxon’ being studied in the Clarion-Clipperton Zone for detailed population genetic study, Paralacydonia sp. This is a likely new species to science, but is abundant in a range of CCZ exploration areas. Image credit: Adrian Glover, Helena Wiklund and Thomas Dahlgren.
Pedro Martinez outlined population studies on ophiuroids being led by the Senckenberg team. Ophiuroids are extremely common in the CCZ, and are often the most recognised metazoan in video transects (Fig.

The species often referred to as Ophiomusium cf. glabrum which is a likely cryptic species complex in CCZ samples, but is present in large numbers and is recognisable (at least to genus) in image surveys. This is a current target taxon for population studies. Image source: (
Uwe Raschka presented an overview of detailed study of a species of harpacticoid copepod, Pseudotachidius bipartitus first described from the Beaufort Sea, Alaska, but recorded from the CCZ in recent surveys. 90 individuals have been sampled, with COI sequences obtained from 57 individuals from across the BGR, IOM, GSR and Ifremer contract areas. Publications on the genetic diversity and cryptic speciation are in preparation.
Pedro Ribeiro presented a preliminary population genetics analysis of a vent mussel species on the Mid-Atlantic Ridge using next-generation sequencing of RAD tag libraries (RADseq). The main purpose of this presentation was to provide an overview of the advantages, as well as methodological and analytical challenges, of using this technique to investigate population genetic structure in the deep sea. RADseq can be more time-efficient compared to other methods, and yield large numbers of single nucleotide polymorphism (SNP) markers, thus holding the potential to uncover levels of genetic structure usually not attainable by traditional methods. However, this technique requires high DNA integrity of samples to construct good quality RAD tag libraries, which in turn are instrumental for sequencing success. Furthermore, discovery and validation of SNP markers can be particularly challenging and may require investing some time for developing the bioinformatics skills necessary to properly explore the data and the array of bioinformatics tools available.
Sergi Taboada presented preliminary data from a new MIDAS-funded study of the detailed population genetics of a relatively newly-discovered and overlooked species of nodule-dwelling sponge (Fig.

A new species of sponge (white object on nodule), first noted on the R/V Melville ABYSSLINE cruise in October 2013, currently in description (Lim et al, National University of Singapore) possibly a new genus related to Hemiasteralla, that is now the subject of detailed investigation by the MIDAS and ABYSSLINE CCZ teams. The scale bar is 1cm. Image credit: Adrian Glover, Thomas Dahlgren, Helena Wiklund.
Session 5: Synthesis of the broader biogeographic context based on morphology and molecules
The purpose of the final data session at the London Workshop was to integrate the new studies of molecular biogeography and connectivity with the broader biogeographic context, in particular data based on morphology. Historically this has been from collected samples in formalin that are unsuited to DNA work; more recently there has been a surge of interest in trying to understand broad ranges of species based on imagery (
Adrian Glover presented an overview of the ‘express Taxonomy’ approach used in the ABYSSLINE project to make raw taxonomic data (species morphology, genetic data and natural history observations) available from recent cruises even before species description is possible. These data are being published by the team in open-access data journals throughout the project. Glover highlighted the importance of morphological data in allowing comparisons with historic collections and species records (see Workshop Recommendations). These comparisons have in many instances supported potentially broad ‘cosmopolitan’ species distributions, but genetic data are in most cases lacking from species type localities (the location of original description). Glover explained how the team had been dealing with this by careful examination of type locality and/or material, and use of open nomenclature abbreviation ‘cf.’ to highlight cosmopolitanism as an untested hypotheses in many instances. An example was presented that links the new DNA data to historical collections and knowledge – that of Nucula profundorum, a protobranch mollusc that is abundant in abyssal samples. Samples of N. profundorum obtained from the ABYSSLINE program were assigned to N. profundorum based on direct morphological comparison with type material held in the NHM collections. Interestingly, on sequencing the specimens, they did not match the published sequences of N. profundorum on GenBank, and it is likely that the sequences on GenBank (recorded at much shallower depths and a long distance from the N. profundorum type locality) are erroneously identified. These matters will be clarified in future publications, but it highlights the importance of improving the quality of data on GenBank and checking the type locality of species that are identified (see Workshop Recommendations).
Daniel Kersken presented an overview of the Senckenberg work on Porifera from the JPI Oceans cruise SO239. The project was focussed not just on DNA work but also on providing an image-based catalogue of the Porifera in the CCZ. Samples were obtained from 15 ROV stations in depths of 1700-5000m in the eastern CCZ from the main SO239 sample areas. A large number of species were recovered, which are the subject of detailed morphological taxonomy coupled with DNA barcoding. Future projects include an attempt at next-generation sequencing and radiocarbon dating of Saccocalyx pedunculatus to determine the age of the sponges, as well as video-based annotation along ROV transects.
Andrew Gooday presented an overview of the broader biogeographic context for the Foraminifera (Fig.

Some well-known deep-water foraminifera that are considered cosmopolitan based on morphology. Images by Andrew Gooday, Aurélie Goineau, ABYSSLINE project.
Gordon Paterson contributed to the session with a talk on the issue of rarity in the deep sea. Understanding species ranges is critical to the assessment of extinction risk, and part of this issue is related to rare species: are they particularly at risk? Paterson showed how almost all deep-sea biodiversity datasets demonstrate a pattern of rarity – that is large numbers of singleton species. Even when modelled against sampling effort, the number of rare species does not seem to decline. Paterson noted that much better definitions of rarity are needed, and is currently working on a meta-analysis of this problem using CCZ data. These show that rare species are the most represented in CCZ samples (in terms of percentages of species) but locally rare (but widely distributed) species also make up a similarly large percentage. So species may be locally rare, but actually widespread and hence common, irrespective of the abundance.
Analysis of megafauna based on imagery
Although not part of the original London Workshop brief, discussions at the workshop also focussed to some extent on information that can be gleaned from imagery surveys. These are increasingly being used by contractors and academic cruises to gain broad-scale information on species abundance and diversity. Recent ISA workshops have recommended that imagery surveys are backed up by sampling (
The megafauna (typically organisms >2 cm in smallest dimension) constitute an important component of the biodiversity in the abyssal CCZ and play a significant role in deep-sea ecosystem function. Knowledge of biogeography and population connectivity are essential for effective environmental management of nodule mining, including the design of marine protected areas.
The study of megafaunal biogeography and connectivity requires collection of information on megafaunal biodiversity via imagery as well as physical samples of organisms, over a range of spatial scales within and beyond the CCZ, followed by morphological and molecular comparisons. The first studies now being published based on imagery data suggest there are many species new to science in the CCZ (
Summary tables and discussion
The final day of the London Workshop (Figs
Summary table produced during the London Workshop on the Biogeography and Connectivity of the Clarion-Clipperton Zone outlining proposed publications during the next three years, designed to facilitate collaboration and complementarity of activities in the region.
| Taxon or Concept Focus | Regional Focus | Lead Institution | Probable Lead Author | Study type | Notes/Collaborations |
| Taxonomy & biogeography | |||||
| Annelida | DISCOL | U Aveiro | Ravara |
Biodiversity and taxonomy |
Potential to collaborate with UK-1 samples (Contact Adrian Glover) |
| Annelida (Data Paper) | UK-1A | NHM London | Wiklund | DNA Taxonomy Catalogue | in prep |
| Bryozoa (Taxonomy) | UK-1A, UK-1 B, OMS, APEI-6 | NHM London | Waeschenbach | Taxonomy | analysis ongoing |
| Cnidaria (Black corals, pennatulids, alcyonaceans) | BGR, GSR, IOM, APEI-3, IFREMER | IMAR - Azores | Ribeiro | Taxonomy and Biogeography | Several papers |
| Cnidaria (Data Paper) | UK-1A | NHM London | Dahlgren | DNA Taxonomy Catalogue | in press |
| Crustacea (Amphipoda) | BGR, IFREMER, APEI-3, IOM, GSR, DISCOL, UK-1 | Belgian Institute | Patel | Biogeography and evolution | Potential to collaborate with UK-1 samples (Contact Tammy Horton) |
| Crustacea (Amphipoda) | UK-1 A, UK-1 B, OMS, APEI-6 | NOC Southampton | Horton | Taxonomy | Potential to collaborate with T Patel (Belgian Institute) |
| Crustacea (Amphipoda) | UK-1A, UK-1 B, OMS, APEI-6 | Senckenberg | Mohrbeck | Taxonomy and population genetics | |
| Crustacea (Cirripedia) | BGR, IFREMER, APEI-3, IOM, GSR, DISCOL, UK-1 | Belgian Institute | Kerkoff | Taxonomy | Potential to collaborate with UK-1, BGR samples (contact P Martinez) |
| Crustacea (Copepoda - benthic) | BGR, IOM, GSR, IFREMER, UK-1 A, UK-1 B, APEI-6 | Senckenberg | Raschka | Biogeography | Potential to collaboate with JC120 (contact S Taboada) |
| Crustacea (Copepoda - benthic) | BGR, IOM, GSR, IFREMER, APEI-3 | Senckenberg | Khodami | Phylogeny | |
| Crustacea (Copepoda) | BGR, IOM, GSR, APEI- 3 | Senckenberg | Khodami | Barcoding, diversity and population genetics | |
| Crustacea (Isopoda) | BGR, IFREMER | Senckenberg | Janessen | DNA Taxonomy Catalogue | |
| Crustacea (Isopoda) | BGR, IOM, GSR, IFREMER, APEI-3, DISCOL, APEI-3, UK-1 A, UK-1 B | Senckenberg | Schnurr | Biodiversity and biogeography | Potential to collaboate with JC120 (contact S Taboada) |
| Crustacea (Isopoda) | BGR, Atlantic | Senckenberg | Kaiser | Taxonony, phylogeny, new genus | |
| Crustacea (Isopoda) | BGR, IOM, GSR, IFREMER, APEI-3, Atlantic | Senckenberg | Schnurr | Phylogeny | |
| Crustacea (Ostracoda) | BGR, IFREMER, APEI-3, IOM, GSR, DISCOL, UK-1 A, UK 1-B, APEI6 | Belgian Institute | Patel | Biogeography and evolution | Potential to collaborate with UK-1, BGR samples (contact P Martinez) |
| Crustacea (Tanaidacea) | BGR, GSR, IOM, APEI-3, IFREMER, Atlantic, APEI-6, UK-1 A, UK-1 B, OMS | U Łódź | Blażewicz | Biodiversity, taxonomy, phylogeny | Potential to collaborate with UK-1, BGR samples (contact P Martinez); GSR (contact A Vanreusel) |
| Echinodermata (Data Paper) | UK-1A, UK-1 B, OMS, APEI-6 | NHM London | Glover | DNA Taxonomy Catalogue | Update to Glover et al 2016 with new taxa |
| Echinodermata (Holuthuroidea) | DISCOL, CCZ | Senckenberg | Martinez | DNA Taxonomy | |
| Echinodermata (Ophiuroidea) | UK-1, APEI | Mus Victoria | O'Hara | Taxonomy | Description of Ophiosphalma sp |
| Ecology - Megafauna (CCZ Seamounts) | BGR, GSR, APEI-3 | IMAR - Azores | Ribeiro | Biogeography | Based on DNA |
| Foraminifera - Xenophyophores | UK-1 A, UK-1 B, OMS, APEI-6, Russian (YMG) | NOC Southampton | Gooday | Biodiversity and biogeography | |
| Foraminifera - Xenophyophores | UK-1 A, UK-1 B, OMS, APEI-6 | NOC Southampton | Gooday | Taxonomy | maybe split into two papers |
| Misc. Taxa (Data Paper, Pycnogonida, Brachiopoda, Sipuncula...) | UK-1A | Uni Research | Dahlgren | DNA Taxonomy Catalogue | in prep |
| Mollusca | DISCOL, UK-1 A, UK-1 B, OMS, APEI-6 | U Aveiro | Rodrigues | Taxonomy | Potential to collaborate with UK-1 samples (Contact Adrian Glover) |
| Mollusca (Data Paper) | UK-1A | NHM London | Wiklund | DNA Taxonomy Catalogue | in prep |
| Nematoda | BGR, IFREMER | Senckenberg | Miljutin | Taxonomy Catalogue | |
| Nematoda | BGR, IOM, GSR, IFREMER, APEI-3, UK-1 A, UK-1 B, OMS, APEI-6 | U Ghent | Macheriotou | Biodiversity and biogeography | Potential to collaborate with UK-1, BGR samples (contact P Martinez) |
| Porifera | BGR, IOM, GSR, IFREMER, APEI-3 | Senckenberg | Kersken | Taxonomy | |
| Porifera | BGR, IOM, GSR, IFREMER, APEI-3 | Senckenberg | Kersken | DNA phylogeography | after first paper above |
| Porifera (Timea / Hemiasterella taxonomy) | OMS | NUS Singapore | Lim | Taxonomy | in prep |
| Tanaidacea | DISCOL | U Aveiro | Esquete | Biodiversity and taxonomy | |
| Methodology | |||||
| Foraminifera | OMS, UK-1 A, UK-1 B | U Geneva | Pawlowski | HTS Methodology | |
| Ecology | |||||
| Echinodermata (Ophiuroidea) | BGR, IOM, GSR, IFREMER, APEI-3 | U Aveiro | Hilário | Life History | |
| Ecology - Macrofauna | GSR | U Ghent | Desmet | Community structure | |
| Ecology - APEI#6 overview | APEI-6 | NOC Southampton | Jones | Ecological overview of the APEI | JC120 and ABYSSLINE |
| Ecology - eDNA | APEI-6 | NOC Southampton | NOC Southampton | eDNA water column and sediment | |
| Ecology - Macrofauna | DISCOL | U Aveiro | Cunha | Community structure | |
| Connectivity | |||||
| Annelida & Crustacea | BGR, IFREMER | Senckenberg | Janssen | Population Genetics | |
| Connectivity - Population Genetics | UK-1A, UK-1 B, OMS, APEI-6 | Uni Research | Dahlgren | Population Genetics | analysis ongoing |
| Connectivity - Workshop Synthesis | CCZ | NHM London | Glover | Synthesis | |
| Echinodermata (Ophiuroidea) | UK-1A, UK-1 B, OMS, APEI-6, APEI-3, GSR, BGR | Senckenberg | Martinez | Population Genetics and Taxonomy | NHM to pass ophiuroid sequences to Senckenberg |
| Foraminifera - Xenophyophores | OMS, UK-1 B, IFREMER | U Geneva | Lejzerowicz | eDNA global synthesis | |
| Porifera (Timea / Hemiasterella populations) | UK-1A, OMS, APEI-6 | NHM London | Taboada | Population Genetics | analysis ongoing |

The London Workshop on the Biogeography and Connectivity of the Clarion-Clipperton Zone discussing data in the Board Room of the Natural History Museum. Image: Adrian Glover.

The London Workshop on the Biogeography and Connectivity of the Clarion-Clipperton Zone on the main steps of the Natural History Museum below the bust of Charles Darwin. Workshop participants: Back row, left to right: Ralph Spickermann, Stefan Brager. Second from back row, left to right: Pedro Ribeiro, Sergi Taboada, Lara Macheriotou, Ann Vanreusel, Magda Blazewicz, Sarah Schnurr, Helena Wiklund. Third from back row, left to right: Andrew Gooday, Daniel Jones, Lenaick Menot, Pedro Martinez, Steffi Kaiser, Ana Colaço, Sahar Khodami, Marina Cunha, Ana Hilario. Third from back row, left to right: Adrian Glover, Paulo Bonifacio, Kirsty McQuaid, Aurélie Goineau, Heiko Stuckas, Thomas Dahlgren. Front row, left to right: Daniel Kersken, Uwe Raschka, Tasnim Patel, Clara Rodrigues, Gordon Paterson, Amber Cobley. Not present for photo: Andrea Waeschenbach and David Billett. Image credit: T Cruise.
The only confirmed cruise plans indicated by the group were for BGR cruises in May 2016 and May 2017. In terms of grant proposals, Adrian Glover highlighted the ongoing work to develop a Strategic Programme Area (SPA) for the Natural Environment Research Council that would focus on deep-sea resource extraction in general, potentially the CCZ could be part of this. At the time of writing, no decisions have yet been made on this proposal. Pedro Martinez indicated that there is interest in developing a second iteration of the JPI Oceans project, but the timeline is not yet known. This may involve technology and mining tests. Some discussion ensued on the idea to develop a broad cross program proposal on taxonomy (see Workshop Recommendations).
Conclusions
Workshop Recommendations
The London Workshop on the Biogeography and Connectivity of the CCZ concluded with a short discussion on Workshop Recommendations, summarised here:
Funding and Planning
- The most critical funding priority for CCZ research on biogeography and connectivity is high-quality taxonomic work that includes species descriptions based on phylogenetic inference, specimen and data archiving to permit future molecular work and equal weight given to all taxonomic groups and size classes
- Taxonomic funding proposals should consider international programs that could deliver taxonomic funding across a range of contractor projects, not just within single ones
- Future international projects on the CCZ must include funding to ensure integrating workshops are fully costed within those projects as they play a vital role in developing collaborations and minimising redundancy of activities
- Future Workshops on CCZ connectivity should be expanded to include non-EU contractors and additional international programs
- Areas of Particular Environmental Interest (APEIs), the majority of which are completely unstudied, should be a priority area for research programs on biogeography and connectivity, helping to deliver an improved regional-level understanding of the CCZ
- Assumptions in the models used for the development of APEIs should be updated as soon as possible with new data with a funded program of research
- The CCZ academic community should be better informed of policy developments and regulatory milestones
- Any biological sampling programs that are taking place in the CCZ must include a funded program of research to work up the samples through to publication and data/specimen archiving
Research
- Connectivity (e.g ecological, population, genetic) should be more precisely defined in CCZ projects and proposals
- Efforts should be undertaken to examine connectivity in rare species, either through NGS approaches or more broad sampling to determine if rare species are common in under-sampled habitats or refugia
- ROV or AUV imagery of megafauna should be improved through (1) collection of voucher materials to confirm conspecificity through DNA sampling, (2) improved resolution, in particular the use of high-quality scientific ROV able to hover and acquire quality imagery and (3) development of regional atlases of megafauna that are available to all contractors and academic programs
- Ongoing and future efforts for the development of automated software tools for video analysis are also fundamental and must be encouraged. Given the huge amount of video data gathered by ROV and AUV surveys, automated methods are essential to be able to produce comprehensive community analyses on a timely manner.
- Working species concepts for new CCZ fauna should be based where possible on phylogenetic species concepts using molecular data in combination with morphological characterization
- New approaches to connectivity study should be developed using new data on larval biology, functional traits (e.g to establish connectivity amongst a range of trait parameters) and physical oceanography (e.g currents and modeled currents)
- New genetic markers and methods to study connectivity should be developed that are appropriate to the fundamental questions posed
Data and Archiving
- Publications on the DNA taxonomy or phylogeography of the Clarion-Clipperton Zone should ensure that raw data in the form of genetic sequences, images, locality information and voucher material access are archived in global online repositories (GenBank, OBIS, GBIF, WoRMS)
- Morphological data (e.g type material in museum collections, taxonomic descriptions and new morphological information) must be used, where possible, alongside molecular data to (1) improve knowledge of putative species ranges, (2) improve taxonomic descriptions that rely on DNA data and (3) check the type material of published species and to use a precautionary approach in estimating species ranges
Funding program
European Union Framework 7 ‘Managing Impacts of Deep Sea Resource Exploitation (MIDAS)’ and Joint Programming Initiative Healthy and Productive Seas and Oceans (JPI Oceans).
Hosting institution
Natural History Museum, London
Author contributions
All authors with the exception of D Amon and C Smith participated in the Workshop. Amon and Smith sent a contribution to the report for inclusion in text form. Adrian Glover led the workshop organisation, chaired the sessions and drafted the manuscript. Amber Cobley took notes at the Workshop. All authors commented and edited the manuscript. The views expressed in this paper are those of the authors and do not necessarily reflect the position of the International Seabed Authority.
Conflicts of interest
None.
References
-
Insights into the abundance and diversity of abyssal megafauna in a polymetallic-nodule region in the eastern Clarion-Clipperton Zone.Scientific Reports6DOI: 10.1038/srep30492
-
Deep-Sea Asteroids: High Genetic Variability in a Stable Environment.Evolution29(2):203‑212. [InEnglish]. DOI: 10.2307/2407208
-
Physically disturbed deep-sea macrofauna in the Peru Basin, southeast Pacific, revisited 7 years after the experimental impact.Deep Sea Research Part II: Topical Studies in Oceanography48(17):3809‑3839. [InEnglish]. DOI: 10.1016/s0967-0645(01)00069-8
-
An Epibenthic Sledge for Operations on Marine Soft Bottom and Bedrock.Marine Technology Society Journal39(2):10‑21. [InEnglish]. DOI: 10.4031/002533205787444015
-
Adaptive response to environmental stability: a unifying concept in paleoecology. Proceedings of the North American Palaeontological Convention.Proc. North American Paleontological Convention,550pp.
-
Patterns of occurence of benthic foraminifera in time and space. In: Donovan S, Paul C (Eds)The Adequacy of the Fossil Record.Chichester,New York,312pp.
-
Abyssal fauna of the UK-1 polymetallic nodule exploration area, Clarion-Clipperton Zone, central Pacific Ocean: Cnidaria.Biodiversity Data Journal4:e9277. DOI: 10.3897/bdj.4.e9277
-
Phylogeography of a pan-Atlantic abyssal protobranch bivalve: implications for evolution in the Deep Atlantic.Molecular Ecology20(4):829‑843. DOI: 10.1111/j.1365-294x.2010.04978.x
-
A chemosynthetic weed: the tubeworm Sclerolinum contortum is a bipolar, cosmopolitan species.BMC Evolutionary Biology15(1):1. DOI: 10.1186/s12862-015-0559-y
-
Project ABYSSLINE Interim Progress Report 2015 - Benthic Biological Studies in the Clarion-Clipperton Zone: Megafaunal and Macrofaunal Biodiversity and Genetics.NHM ConsultingSBU13002/15.
-
Morphology, reproductive biology and genetic structure of the whale-fall and hydrothermal vent specialist, Bathykurila guaymasensisPettibone, 1989 (Annelida: Polynoidae).Marine Ecology26:223‑234. DOI: 10.1111/j.1439-0485.2005.00060.x
-
An End-to-End DNA Taxonomy Methodology for Benthic Biodiversity Survey in the Clarion-Clipperton Zone, Central Pacific Abyss.Journal of Marine Science and Engineering4(1):2. DOI: 10.3390/jmse4010002
-
Abyssal fauna of the UK-1 polymetallic nodule exploration claim, Clarion-Clipperton Zone, central Pacific Ocean: Echinodermata.Biodiversity Data Journal4:e7251. DOI: 10.3897/bdj.4.e7251
-
Benthic Foraminiferal Biogeography: Controls on Global Distribution Patterns in Deep-Water Settings.Annual Review of Marine Science4(1):237‑262. DOI: 10.1146/annurev-marine-120709-142737
-
Species diversity in the cryptic abyssal holothurian Psychropotes longicauda (Echinodermata).Deep Sea Research Part II: Topical Studies in Oceanographyin pressDOI: 10.1016/j.dsr2.2016.04.003
-
Environmental Management Plan for the Clarion-Clipperton Zone. https://www.isa.org.jm/documents/isba17ltc7
-
Workshop on taxonomic methods and standardization of megafauna in the Clarion-Clipperton Fracture Zone. https://www.isa.org.jm/documents/briefing-paper-0214
-
Workshop on taxonomic methods and standardization of macrofauna in the Clarion-Clipperton Fracture Zone. https://www.isa.org.jm/workshop/workshop-taxonomic-methods-and-standardization-macrofauna-clarion-clipperton-fracture-zone
-
Workshop on taxonomic methods and standardization of meiofauna in the Clarion-Clipperton Fracture Zone. https://www.isa.org.jm/workshop/workshop-taxonomic-methods-and-standardization-meiofauna-clarion-clipperton-zone-dec-2015
-
A Reverse Taxonomic Approach to Assess Macrofaunal Distribution Patterns in Abyssal Pacific Polymetallic Nodule Fields.PLOS ONE10(2):e0117790. DOI: 10.1371/journal.pone.0117790
-
Short Cruise Report R.V Sonne Cruise SO-239. https://www.ldf.uni-hamburg.de/sonne/wochenberichte/wochenberichte-sonne/so239/so239-scr.pdf
-
Global biodiversity indices from the Ocean Biogeographic Information System. Intergovernmental Oceanographic Commission of UNESCO. http://www.iobis.org
-
Current status of manganese nodule exploration in the German license area.Proceedings of the Ninth (2011) ISOPE Ocean Mining Symposium: Maui, Hawaii, USA, June 19-24. Cupertino, Calif. : International Society of Offshore and Polar Engineers (ISOPE).168-173pp. [ISBN978-1-880653-95-1].
-
Fully automated image segmentation for benthic resource assessment of poly-metallic nodules.Methods in Oceanography15-16:78‑89. DOI: 10.1016/j.mio.2016.04.002
-
Review of Implementation of the Environmental Management Plan for the Clarion-Clipperton Zone.Seascape Consultants Report2014.
-
Preservation Reference Areas for Nodule Mining in the Clarion-Clipperton Zone: Rationale and Recommendations to the International Seabed Authority. International Seabed Authority.http://www.soest.hawaii.edu/oceanography/faculty/csmith/MPA_webpage/documents/Smith%20et%20al.%20-%20Recommendations%20to%20the%20ISA%20for%20design%20of%20PRAs%20in%20the%20CCZ%20-%202-2008.pdf.
-
Biodiversity, species ranges, and gene flow in the abyssal Pacific nodule province: predicting and managing the impacts of deep seabed mining: report. http://www.isa.org.jm/files/documents/EN/Pubs/TechStudy3.pdf
-
From principles to practice: a spatial approach to systematic conservation planning in the deep sea.Proceedings of the Royal Society B: Biological Sciences280(1773):20131684‑20131684. DOI: 10.1098/rspb.2013.1684